
replikasi dari DNA (asam deoksiribonukleat) adalah untuk menyalin genom, yaitu, semua informasi genetik dalam DNA dari suatu organisme untuk menghasilkan dua salinan identik. Genom memiliki informasi yang diperlukan untuk membangun organisme yang lengkap.
Sebelum pembelahan sel, terjadi replikasi DNA. Melalui meiosis, gamet diproduksi untuk reproduksi seksual. Melalui mitosis, penggantian sel (misalnya, kulit dan darah) dan perkembangan (misalnya, jaringan dan organ) terjadi.
Sumber: I, Madprime [CC0]
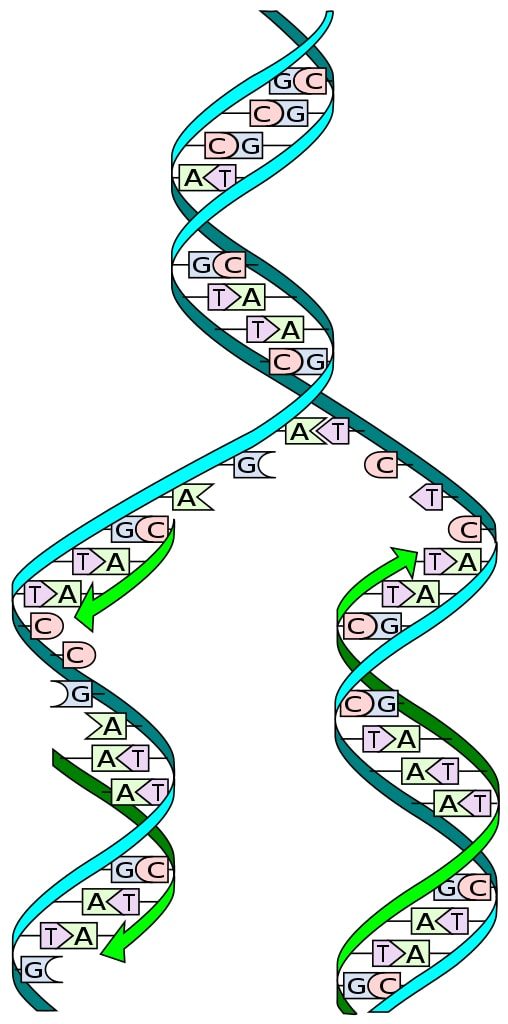
Mengetahui struktur DNA memungkinkan kita untuk memahami bagaimana replikasinya terjadi. Struktur DNA terdiri dari heliks ganda, terdiri dari dua rantai antiparalel dari nukleotida berturut-turut, yang basa nitrogennya saling melengkapi dengan cara tertentu.
Selama replikasi, setiap untai untai ganda DNA bertindak sebagai cetakan untuk biosintesis untai baru. Dua rantai yang baru disintesis memiliki basa yang saling melengkapi dengan basa rantai template: adenin (A) dengan timin (T), dan sitosin (C) dengan guanin (G).
Berbagai enzim dan protein terlibat dalam replikasi DNA. Misalnya, membuka heliks ganda DNA, menjaga DNA tetap terbuka, dan menambahkan deoxyribonucleosides-5′-triphosphate (dNTP) untuk membentuk untai baru.
Indeks artikel
Replikasi DNA bersifat semi-konservatif
Berdasarkan struktur DNA, Watson dan Crick mengusulkan agar replikasi DNA terjadi secara semi-konservatif. Hal ini ditunjukkan oleh Meselson dan Stahl dengan memberi label DNA Escherichia coli dengan isotop nitrogen berat, 15 N, mengikuti pola distribusi dalam media kultur dengan nitrogen ringan, 14 N selama beberapa generasi .
Meselson dan Stahl menemukan bahwa, pada generasi pertama, dua molekul DNA anak memiliki setiap molekul yang diberi label dengan rantai dengan isotop berat nitrogen dan satu lagi dengan isotop ringan. Berbeda dengan molekul DNA induk, yang memiliki kedua untai berlabel isotop berat, 15 N.
Pada generasi kedua, 50% molekul DNA seperti generasi pertama, dan 50% lainnya hanya memiliki nitrogen ringan. Interpretasi dari hasil ini adalah bahwa anak heliks ganda memiliki rantai induk (yang berfungsi sebagai template) dan rantai baru.
Mekanisme replikasi semi-konservatif melibatkan pemisahan untai DNA dan pasangan basa komplementer melalui pasangan nukleotida berturut-turut, menghasilkan dua heliks ganda putri.
Replikasi baterai
Inisiasi replikasi DNA pada bakteri
DNA bakteri terdiri dari kromosom melingkar dan hanya memiliki satu tempat asal replikasi. Dari situs ini, biosintesis dua rantai anak terjadi secara dua arah, membentuk dua garpu replikasi yang bergerak berlawanan arah dengan asalnya. Pada akhirnya, jepit rambut bertemu, menyelesaikan replikasi.
Replikasi dimulai dengan pengikatan protein DNA ke situs asal. Protein ini pada gilirannya membentuk kompleks. Kemudian protein HU dan IHF, antara lain, bergabung bersama, yang bersama-sama melipat DNA, menyebabkan pemisahan dua untai DNA di daerah yang kaya akan timin dan adenin.
Selanjutnya, protein DNaC mengikat, yang menyebabkan helikase DNA untuk mengikat. Mereka membantu melepaskan DNA dan memutuskan ikatan hidrogen, yang terbentuk di antara pasangan basa. Jadi kedua rantai tersebut dipisahkan lebih lanjut, membentuk dua rantai sederhana.
Topoisomerase II, atau DNA girase, bergerak di depan DNA helicase, mengurangi superkoil positif. Protein pengikat DNA untai tunggal (SSB) menjaga untaian DNA tetap terpisah. Dengan demikian, biosintesis rantai anak dapat dimulai.
Biosintesis untai DNA anak pada bakteri
Enzim primase bertanggung jawab untuk sintesis rantai RNA pendek yang disebut primer, yang 10 sampai 15 nukleotida panjang. DNA polimerase mulai menambahkan deoxynucleosides 5′-trifosfat (dNTP) ke ujung 3′-OH dari gula primer, setelah untai terus tumbuh dari ujung yang sama.
Karena untai DNA bersifat antiparalel, satu primer disintesis pada untai pemimpin dan banyak primer pada untai tertinggal. Karena itu, biosintesis rantai tertunda terputus-putus. Meskipun untai DNA antiparalel, garpu replikasi hanya bergerak dalam satu arah.
DNA polimerase bertanggung jawab untuk pembentukan ikatan kovalen antara nukleotida yang berdekatan dari rantai yang baru disintesis, dalam arah 5’®3 . Dalam E. coli , ada lima DNA polimerase: DNA polimerase I dan III melakukan replikasi DNA; dan DNA polimerase II, IV dan V bertanggung jawab untuk memperbaiki dan mereplikasi DNA yang rusak.
Sebagian besar replikasi dilakukan oleh DNA polimerase III, yang merupakan holoenzim yang memiliki 10 subunit berbeda dengan berbagai fungsi dalam replikasi DNA. Misalnya, subunit alfa bertanggung jawab untuk membuat hubungan antara nukleotida.
Kompleks enzim bertanggung jawab untuk replikasi DNA pada bakteri
DNA helikase dan primase bergabung bersama untuk membentuk kompleks yang disebut primosom. Ini bergerak di sepanjang DNA, bertindak secara terkoordinasi untuk memisahkan dua untai induk, mensintesis primer setiap interval tertentu pada untai tertunda.
Primosom secara fisik mengikat DNA polimerase III, dan membentuk replisome. Dua DNA polimerase III bertanggung jawab untuk mereplikasi DNA pemandu dan rantai tertunda. Sehubungan dengan DNA polimerase III, untai terbelakang membentuk loop ke luar, yang memungkinkan penambahan nukleotida ke untai ini terjadi dalam arah yang sama dengan untai pemimpin.
Penambahan nukleotida ke rantai pemandu terus menerus. Sedangkan pada delay bersifat diskontinyu. Fragmen 150 nukleotida panjang terbentuk, disebut fragmen Okazaki.
Aktivitas eksonuklease DNA polimerase I 5 -> 3 bertanggung jawab untuk menghilangkan primer dan mengisi, menambahkan nukleotida. Enzim ligase menutup celah di antara fragmen. Replikasi berakhir ketika dua kait replikasi bertemu dalam urutan terminasi.
Protein Tus mengikat urutan terminasi, menghentikan pergerakan garpu replikasi. Topoisomerase II memungkinkan pemisahan dua kromosom.
Deoksiribonukleotida trifosfat digunakan oleh DNA polimerase
Deoxynucleoside triphosphate (dNTP) mengandung tiga gugus fosfat yang terikat pada 5 karbon deoksiribosa. dNTPs (dATP, dTTP, dGTP dan dCTP) mengikat rantai template mengikuti aturan AT / GC.
DNA polimerase mengkatalisis reaksi berikut: gugus 3 hidroksil (–OH) dari nukleotida untai yang tumbuh bereaksi dengan alfa fosfat dari dNTP yang masuk, melepaskan pirofosfat anorganik (PPi). Hidrolisis PPi menghasilkan energi untuk pembentukan ikatan kovalen, atau ikatan fosfodiester, antara nukleotida rantai yang sedang tumbuh.
Mekanisme yang memastikan ketepatan replikasi DNA
Selama replikasi DNA, DNA polimerase III membuat kesalahan sebanyak 100 juta nukleotida. Meskipun kemungkinan kesalahan sangat rendah, ada mekanisme yang memastikan ketepatan dalam replikasi DNA. Mekanisme ini adalah:
1) Stabilitas dalam pasangan basa. Energi ikatan hidrogen antara AT / GC lebih tinggi daripada pasangan basa yang salah.
2) Struktur situs aktif DNA polimerase. DNA polimerase secara istimewa mengkatalisis sambungan nukleotida dengan basa yang benar pada untai yang berlawanan. Pasangan basa yang buruk menghasilkan distorsi heliks ganda DNA, yang mencegah nukleotida yang salah menempati situs aktif enzim.
3) Tes membaca. DNA polimerase mengidentifikasi nukleotida salah yang tergabung dan menghilangkannya dari untai anak. Aktivitas eksonuklease DNA polimerase memutus ikatan fosfodiester antara nukleotida pada ujung 3 untai baru.
Replikasi DNA pada eukariota
Tidak seperti replikasi pada prokariota, di mana replikasi dimulai di satu tempat, replikasi pada eukariota dimulai di banyak tempat asal dan garpu replikasi bergerak dua arah. Kemudian, semua jepit rambut replikasi sekering, membentuk dua kromatid kakak bergabung di sentromer.
Eukariota memiliki banyak jenis DNA polimerase, yang namanya menggunakan huruf Yunani. DNA polimerase α membentuk kompleks dengan Primase. Kompleks ini mensintesis primer pendek yang terdiri dari 10 nukleotida RNA diikuti oleh 20 hingga 30 nukleotida DNA.
Berikutnya, ε atau δ DNA polimerase mengkatalisis pemanjangan untai putri dari primer. DNA polimerase ε terlibat dalam sintesis rantai pemimpin, sedangkan DNA polimerase δ mensintesis rantai terbelakang.
DNA polimerase δ memanjang fragmen Okazaki di sebelah kiri sampai mencapai primer RNA di sebelah kanan, memproduksi flap singkat primer. Tidak seperti prokariota, di mana DNA polimerase menghilangkan primer, pada eukariota, enzim Flap endonuklease menghilangkan primer RNA.
Ligase DNA kemudian menyegel fragmen DNA yang berdekatan. Terminasi replikasi terjadi dengan disosiasi protein dari garpu replikasi.
Replikasi DNA pada eukariota dan siklus sel
Replikasi pada eukariota terjadi pada fase S dari siklus sel. Molekul DNA yang direplikasi disekresikan ke dalam dua sel anak selama mitosis. Fase G1 dan G2 memisahkan fase S dan mitosis. Kemajuan melalui setiap fase siklus sel sangat diatur oleh kinase, fosfatase, dan protease.
Pada fase G1 dari siklus sel, kompleks pengenalan asal (OCR) berikatan dengan situs asal. Ini menginduksi pengikatan helikase MCM dan protein lain, seperti Cdc6 dan Cdt1, untuk membentuk kompleks pra-replikasi (preRC). Helikase MCM mengikat rantai pemandu.
Pada fase S, preRC menjadi situs replikasi aktif. Protein OCR, Cdc6, dan Cdt1 dilepaskan, dan helikase MCM bergerak dalam arah 3 hingga 5 . Setelah replikasi selesai, itu akan dimulai kembali di siklus sel berikutnya.
Replikasi ujung kromosom pada eukariota
Ujung kromosom dikenal sebagai telomer, yang terdiri dari urutan tandem berulang, dan daerah 3 yang menonjol, dengan panjang 12 hingga 16 nukleotida.
DNA polimerase tidak dapat mereplikasi ujung 3 dari untai DNA. Ini karena DNA polimerase hanya dapat mensintesis DNA dalam arah 5′-3′, dan hanya dapat memperpanjang untaian yang sudah ada sebelumnya, tanpa dapat mensintesis primer di wilayah ini. Akibatnya, telomer memendek dengan setiap putaran replikasi.
enzim telomerase mencegah pemendekan telomere. Telomerase adalah enzim yang memiliki protein dan subunit RNA (TERC). mengikat kedua untuk urutan berulang dari DNA, dan memungkinkan telomerase untuk mengikat ujung 3 ‘dari telomer.
Urutan RNA di belakang situs persimpangan berfungsi sebagai templat untuk sintesis urutan enam nukleotida (polimerisasi) di ujung untai DNA. Pemanjangan telomer dikatalisis oleh subunit telomerase, yang disebut telomerase reverse transcriptase (TERT).
Setelah polimerisasi, terjadi translokasi, yang terdiri dari pergerakan telomerase ke ujung baru rantai DNA, bergabung dengan enam nukleotida lainnya sampai akhir.
Fungsi DNA polimerase lain pada eukariota
DNA polimerase β memainkan peran penting dalam menghilangkan basa yang salah dari DNA, tetapi tidak terlibat dalam replikasi DNA.
Banyak DNA polimerase yang ditemukan termasuk dalam kelompok polimerase “translesi-replikasi”. Polimerase ini bertanggung jawab untuk mensintesis untai komplementer di wilayah DNA yang rusak.
Ada beberapa jenis polimerase “replikasi-translesi”. Misalnya, DNA polimerase η dapat mereplikasi pada timin dimer, yang diproduksi oleh sinar UV.
Replikasi DNA pada archaebacteria
Replikasi DNA Archaebacterial mirip dengan yang ada pada eukariota. Hal ini disebabkan oleh hal-hal berikut: 1) protein yang berpartisipasi dalam replikasi lebih mirip dengan eukariota daripada prokariota; dan 2) meskipun hanya ada satu tempat replikasi seperti pada prokariota, urutannya mirip dengan tempat asal eukariota.
Kesamaan dalam replikasi antara Archea dan eukariota mendukung gagasan bahwa kedua kelompok secara filogenetik lebih terkait satu sama lain daripada dengan prokariota.
Referensi
- Brooker, RJ 2018. Analisis dan prinsip genetika. McGraw-Hill, New York.
- Hartwell, LH, Goldberg, ML, Fischer, JA, Hood, L. 2018. Genetika – dari gen ke genom. McGraw-Hill, New York.
- Kušić-Tišma, J. 2011. Aspek fundamental dari replikasi DNA. Akses InTech Open, Kroasia.
- Lewis, R., 2015. Konsep dan aplikasi genetika manusia. McGraw-Hill, New York.
- Pierce, BA 2005. Genetika – pendekatan konseptual. WH Freeman, New York.
