
enolase adalah enzim yang bertanggung jawab untuk melakukan konversi dari D-2-fosfogliserat (2PGA) phosphoenolpyruvate (PEP) di glikolisis dan glukoneogenesis reaksi sebaliknya, dua jalur metabolisme adalah bagian dari metabolisme energi sel.
Keputusan untuk mengkatalisis reaksi ini dalam satu arah atau yang lain tergantung pada akses sel ke glukosa. Artinya, dari kebutuhan Anda harus menyesuaikan metabolisme Anda dengan degradasi atau sintesis untuk mendapatkan energi. Sangat diperlukan untuk realisasi proses vital mereka.
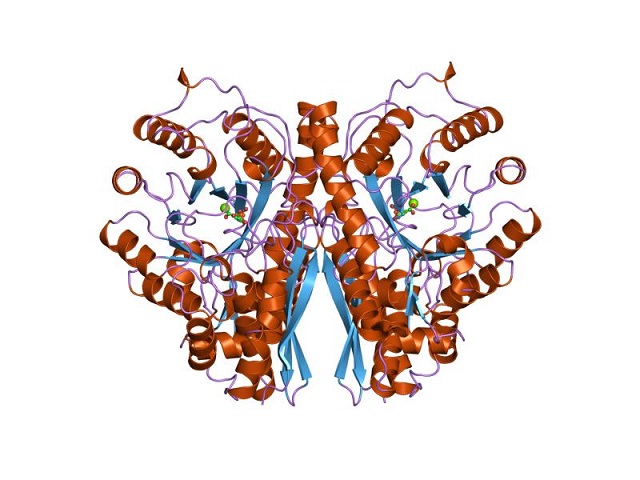
Struktur tiga dimensi Enolase. Oleh Jawahar Swaminathan dan staf MSD di European Bioinformatics Institute [Domain publik (https://creativecommons.org/licenses/by-sa/4.0)], dari Wikimedia Commons.
Karena kedua jalur metabolisme tersebut termasuk dalam pusat pohon metabolisme pusat makhluk hidup, tidak mengherankan bahwa urutan asam amino protein ini disimpan dalam archaea, bakteri, dan eukariota. Dan karena itu ia memiliki sifat katalitik yang serupa.
Lokalisasi enolase dalam sel terbatas pada sitosol, kompartemen di mana kedua glikolisis (juga disebut glikolisis) dan glukoneogenesis berlangsung di sebagian besar organisme.
Namun, itu juga telah terdeteksi di kompartemen sel lain seperti membran plasma dari banyak patogen dan sel kanker. Di sana, tampaknya terlibat dengan fasilitasi proses penyebaran sel, fungsi yang sama sekali berbeda dari fungsi klasiknya.
Enzim yang mampu melakukan lebih dari satu fungsi, seperti enolase, dikenal sebagai enzim moonlighting.
Indeks artikel
Struktur
Struktur kuaterner dari enolase yang terikat atau tidak dengan ligannya telah ditentukan pada sejumlah besar individu prokariotik dan eukariotik.
Setiap monomer memiliki dua domain: domain terminal amino kecil dan domain terminal karboksil yang lebih besar. Domain N-terminal terdiri dari tiga heliks dan empat lembar. Sedangkan C-terminal terdiri dari delapan -sheet yang berselang-seling, membentuk -barrel yang dikelilingi oleh delapan -helices.
Selanjutnya, dua situs pengikatan untuk kation divalen ditemukan pada setiap monomer yang disebut “situs konformasi” dan “situs katalitik”. Yang pertama tidak terlalu selektif dan dapat mengikat berbagai macam kation divalen tanpa adanya substrat.
Sedangkan, yang kedua mengikat ion setelah substrat terikat pada enzim. Pengikatan ion ke kedua situs sangat penting untuk reaksi untuk melanjutkan.
Akhirnya, penting untuk disebutkan bahwa dalam homodimer, monomer-monomer tersebut digabungkan dengan mempertahankan orientasi paralel. Oleh karena itu, situs aktif terbatas pada wilayah tengah yang dibentuk oleh persimpangan tersebut.
Namun, hanya residu dari salah satu dari dua monomer yang berpartisipasi dalam katalisis. Ini menjelaskan kemampuan monomer untuk melakukan reaksi di bawah kondisi eksperimental.
Mekanisme aksi
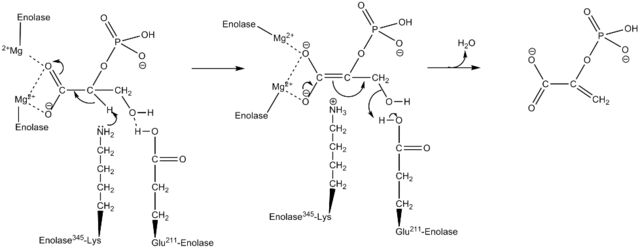
Mekanisme kerja yang digunakan oleh enzim Enolase. Oleh Kthompson08 di Wikipedia bahasa Inggris [Domain publik (https://creativecommons.org/licenses/by-sa/4.0)], dari Wikimedia Commons.
Studi struktural, serta studi yang memungkinkan untuk menentukan karakteristik kinetik dan fisikokimia enolase, telah memungkinkan untuk memahami mekanisme kerjanya.
Cara enzim mengkatalisis reaksi cukup menarik. Terlepas dari kenyataan bahwa satu substrat terlibat, mekanisme berurutan yang telah diusulkan adalah apa yang telah diusulkan.
Ini dimulai dengan pengikatan ion Mg2 + ke situs konformasi salah satu monomer. Ini berlanjut dengan pengikatan substrat ke situs aktif diikuti dengan pengikatan ion kedua ke situs katalitik dan diakhiri dengan pelepasan produk segera setelah reaksi dilakukan. Pada titik ini, Mg2+ tetap melekat pada situs konformasi.
Sejalan dengan itu, untuk mendorong reaksi, enzim pertama-tama memediasi pembentukan zat antara karbanion, menghilangkan proton dari karbon 2 dari 2PGA. Hal ini berkat aksi residu asam amino basa.
Secara berurutan, penghilangan hidroksil karbon 3 terjadi oleh aksi residu asam dari enzim. Pada titik ini, penyatuan kedua karbon dilakukan melalui ikatan rangkap yang membentuk PEP. Dengan cara ini, reaksi dihentikan.
Fitur
Banyak enzim yang dipelajari sejauh ini mampu melakukan berbagai macam fungsi yang tidak terkait dengan “fungsi klasik” mereka di kompartemen sel yang berbeda. Enzim-enzim ini telah disebut sebagai enzim “penerangan bulan”.
Dalam pengertian ini, enolase dapat dianggap sebagai enzim moonlighting, karena banyak fungsi yang bertentangan dengan fungsi klasiknya telah dikaitkan dengannya hingga saat ini baik pada bakteri maupun eukariota.
Beberapa fungsi tersebut adalah sebagai berikut:
– Berpartisipasi dalam pemeliharaan bentuk sel serta lalu lintas vesikular dengan berinteraksi dengan protein sitoskeleton.
– Dalam inti sel mamalia, ia bertindak sebagai faktor transkripsi yang mengatur ekspresi gen yang terkait dengan proliferasi sel. Ini bekerja sama dalam menjaga stabilitas mRNA dalam degradosom pada bakteri.
– Pada patogen, seperti Streptococcus pneumoniae dan Trypanosoma cruzi, tampaknya berperan sebagai faktor virulensi yang penting.
– Juga telah ditemukan bahwa pada Streptococcus pyogenes, enolase diekskresikan ke lingkungan ekstraseluler, memfasilitasi degradasi jaringan dan penghindaran sistem imun pada pejamu.
– Diekspresikan pada permukaan sel tumor, meningkatkan metastasis.
Eolase dan hubungannya dengan mekanisme penyebaran sel
Banyak patogen, serta sel tumor, mengekspresikan dalam membran mereka atau mengeluarkan protease yang mampu mendegradasi protein matriks ekstraseluler ke dalam lingkungan ekstraseluler.
Kemampuan ini memungkinkan sel-sel ini menembus jaringan dan menyebar dengan cepat ke seluruh organisme inang. Mendukung dengan cara ini penghindaran dari sistem kekebalan dan oleh karena itu, pembentukan infeksi.
Meskipun enolase tidak memiliki aktivitas protease, ia berpartisipasi dalam proses penyebaran banyak patogen di inangnya serta sel tumor selama metastasis.
Ini dicapai berkat fakta bahwa itu diekspresikan pada permukaan sel-sel ini dengan berfungsi sebagai reseptor plasminogen. Yang terakhir adalah zymogen dari protease serin yang dikenal sebagai plasmin yang merupakan bagian dari sistem fibrinolitik dan bekerja dengan mendegradasi protein matriks ekstraseluler.
Dengan demikian, enolase yang diekspresikan permukaan adalah strategi yang diperoleh sel-sel ini untuk membangun infeksi dan menyebar dengan sukses.
Strategi ini terdiri dari dua proses:
– Penghindaran dari sistem kekebalan inang. Karena sel-sel ini dilapisi dengan protein inangnya sendiri, mereka diabaikan oleh sel-sel sistem kekebalan yang mengenali protein non-diri yang terkait dengan patogen.
– Diseminasi pasca aktivasi plasminogen dalam plasmin. Yang partisipasinya dalam degradasi protein matriks ekstraseluler, kemudian memfasilitasi penyebaran yang cepat dan efektif.
Referensi
- Avilan L, Gualdron-Lopez M, Quiñones W, González-González L, Hannaert V, Michels PAA, Concepción JL. Enolase: pemain kunci dalam metabolisme dan kemungkinan faktor virulensi parasit trypanosomatid-perspektif untuk penggunaannya sebagai target terapi. Penelitian Enzim. 2011 jilid. Pasal ID932549, 14 halaman.
- Bhowmick I, Kumar N, Sharma S, Coppens I, Jarori GK, Plasmodium falciparum enolase: ekspresi khusus tahap dan lokalisasi sub-seluler. Jurnal Malaria. 2009; 8 (1). pasal 179.
- Hari I, Peshavaria M, Quinn GB, Jam molekuler diferensial dalam evolusi isoprotein enolase. Jurnal Evolusi Molekuler. 1993; 36 (6): 599-601.
- de la Torre-Escudero E, Manzano-Román R, Pérez-Sánchez R, Siles-Lucas M, Oleaga A. Kloning dan karakterisasi enolase terkait permukaan pengikat plasminogen dari Schistosoma bovis. Parasitologi Hewan. 2010; 173: 73-84.
- Dinovo EC, Boyer PD. Probe isotop dari mekanisme reaksi enolase. Nilai tukar isotop awal dan keseimbangan: efek isotop primer dan sekunder. J Biol Kimia 1971; 246 (14): 4586-4593.
- Kaberdin VR, Lin-Chao S, Mengungkap peran baru untuk komponen kecil dari degradosom RNA E. coli. Biologi RNA. 2009; 6 (4): 402-405.
- Keller A, Peltzer J, Carpentier G. Interaksi enolase isoform dengan tubulin dan mikrotubulus selama miogenesis. Biochimica et Biophysica Acta.2007; 1770 (6): 919-926.
- Lung J, Liu KJ, Chang JY, Leu SJ, Shih NY. MBP-1 secara efisien dikodekan oleh transkrip alternatif gen ENO1 tetapi setelah translasi diatur oleh pergantian protein yang bergantung pada proteasom. Jurnal FEBS. 2010; 277 (20): 4308-4321.
- Pancholi V. Multifungsi -enolase: perannya dalam penyakit. Ilmu Hayati Seluler dan Molekuler. 2001; 58 (7): 902-920.
- Poyner RR, Cleland WW, Reed GH. Peran ion logam dalam katalisis oleh enolase. Mekanisme kinetik yang teratur untuk enzim substrat tunggal. Biokimia. 2001; 40: 9008-8017.
- Segovia-Gamboa NC, Chávez-Munguía B, Medina-Flores A, Entamoeba menyerang , proses encystation dan enolase. Parasitologi Eksperimental. 2010; 125 (2): 63-69.
- Tanaka M, Sugisaki K, Nakashima K, Beralih di tingkat mRNA yang dapat diterjemahkan untuk isozim enolase selama pengembangan otot rangka ayam. Komunikasi penelitian biokimia dan biofisika. 1985; 133 (3): 868-872.
