
rekombinasi genetik adalah proses dimana molekul fragmen asam nukleat ditukar menghasilkan molekul baru. Ini sangat umum dalam DNA , tetapi RNA juga merupakan substrat untuk rekombinasi. Rekombinasi adalah, setelah mutasi, sumber yang paling penting dari generasi variabilitas genetik.
DNA berpartisipasi dalam proses biokimia yang berbeda. Selama replikasi, ia berfungsi sebagai cetakan untuk generasi dua molekul DNA baru. Dalam transkripsi, memungkinkan untuk menghasilkan molekul RNA dari daerah tertentu yang dikendalikan oleh promotor.
Langkah-langkah umum rekombinasi DNA. Juergen Bode [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], melalui Wikimedia Commons
Tapi selain itu, DNA juga mampu bertukar fragmen. Melalui proses ini dihasilkan kombinasi-kombinasi baru yang bukan merupakan produk dari dua proses sebelumnya, maupun dari fertilisasi.
Semua proses rekombinasi melibatkan pemutusan dan penggabungan molekul DNA yang berpartisipasi dalam proses tersebut. Mekanisme ini bervariasi tergantung pada substrat rekombinasi, enzim yang berpartisipasi dalam proses, dan mekanisme pelaksanaannya.
Rekombinasi umumnya tergantung pada keberadaan daerah komplementer, serupa (jika tidak identik), atau homolog antara molekul yang bergabung kembali. Dalam hal molekul bergabung kembali dalam proses yang tidak dipandu oleh homologi, rekombinasi dikatakan non-homolog.
Jika homologi melibatkan wilayah yang sangat pendek yang ada di kedua molekul, rekombinasi dikatakan spesifik lokasi.
Indeks artikel
Definisi
Apa yang kita sebut homologi dalam rekombinasi tidak selalu mengacu pada asal evolusi molekul yang berpartisipasi. Sebaliknya, kita berbicara tentang tingkat kesamaan dalam urutan nukleotida.
Rekombinasi non-reparatif, misalnya, terjadi pada eukariota selama proses meiosis. Tidak diragukan lagi, tidak ada homologi yang lebih besar daripada antara pasangan kromosom dalam sel yang sama.
Itulah sebabnya mereka disebut kromosom homolog. Namun, ada kasus di mana DNA suatu sel bertukar materi dengan DNA asing. DNA ini harus sangat mirip untuk bergabung kembali, tetapi mereka tidak harus memiliki nenek moyang yang sama (homologi) untuk mencapai hal ini.
Chiasme dan crossover
Tempat perlekatan dan pertukaran antara dua molekul DNA disebut kiasma, dan prosesnya sendiri disebut ikatan silang. Selama crossover, pertukaran pita diverifikasi antara DNA yang berpartisipasi.
Ini menghasilkan kointegrasi, yang merupakan dua molekul DNA yang secara fisik bergabung menjadi satu. Ketika kointegrasi “terputus”, dua molekul dihasilkan, umumnya berubah (rekombinan).
Untuk “memecahkan”, dalam konteks rekombinasi, adalah untuk memisahkan molekul DNA komponen dari kointegrasi.
Jenis rekombinasi genetik
-Rekombinasi khusus situs
Dalam rekombinasi spesifik lokasi, dua molekul DNA, umumnya tidak homolog, memiliki urutan pendek yang sama untuk keduanya. Urutan ini adalah target dari enzim splicing tertentu.
Enzim, yang mampu mengenali urutan ini dan bukan yang lain, memotongnya di tempat tertentu di kedua molekul. Dengan bantuan beberapa faktor lain, ia menukar pita DNA dari dua molekul yang berpartisipasi dan membentuk kointegrasi.
Escherichia coli dan lambda
Ini adalah dasar untuk pembentukan kointegrasi antara genom bakteri Escherichia coli dan bakteriofag lambda. Bakteriofag adalah virus yang menginfeksi bakteri.
Pembentukan kointegrasi ini dilakukan oleh enzim yang dikodekan dalam genom virus: lambda integrase. Ia mengenali urutan umum yang disebut attP dalam genom sirkular virus, dan attB dalam genom bakteri.
Dengan memotong kedua urutan di kedua molekul, itu menghasilkan segmen bebas, menukar pita, dan menggabungkan dua genom. Lingkaran yang lebih besar atau terkointegrasi kemudian terbentuk.
Dalam kointegrasi, genom virus secara pasif dibawa oleh genom bakteri, yang dengannya ia bereplikasi. Dalam keadaan ini dikatakan bahwa virus berada dalam keadaan provirus, dan bakteri bersifat lisogenik untuk itu.
Proses sebaliknya, yaitu penyelesaian kointegrasi, dapat memakan waktu beberapa generasi – atau bahkan tidak terjadi. Namun, melakukannya secara enzimatis dimediasi oleh protein lain yang dikodekan oleh genom virus yang disebut excisionase. Ketika ini terjadi, virus memisahkan diri dari kointegrasi, mengaktifkan kembali dan menyebabkan lisis sel.
-Rekombinasi homolog
Rekombinasi umum
Rekombinasi homolog terjadi antara molekul DNA yang memiliki setidaknya sekitar 40 nukleotida dengan kemiripan yang lengkap atau hampir sempurna. Untuk melakukan proses rekombinasi, setidaknya satu endonuklease harus terlibat.
Endonuklease adalah enzim yang membuat pemotongan internal DNA. Beberapa melakukannya untuk melanjutkan untuk mendegradasi DNA. Lainnya, seperti dalam kasus rekombinasi, melakukannya untuk menghasilkan penyok pada DNA.
Nick unik ini memungkinkan pemrosesan DNA pita tunggal dengan ujung bebas. Ujung bebas ini, diorientasikan oleh rekombinase, memungkinkan pita tunggal menyerang DNA ganda, menggantikan pita tetap yang identik dengannya.
Ini adalah titik persilangan antara molekul DNA donor (“penyerbu”) dan reseptor lain.
Enzim (rekombinase) yang melakukan invasi dan proses pertukaran pita pada Escherichia coli disebut RecA. Ada protein homolog lain dalam prokariota, seperti RadA di archaea. Pada eukariota, enzim yang setara disebut RAD51.
Setelah pita invasif menggantikan residen, ia berinteraksi dengan pita yang dibiarkan sederhana dalam molekul donor. Kedua situs disegel oleh aksi ligase.
Sekarang kita memiliki DNA pita hibrida (pita donor dan pita penerima, dari asal yang berbeda) diapit oleh DNA donor dan DNA penerima. Titik persilangan (chiasmata) bergerak ke dua arah paling sedikit 200 bp.
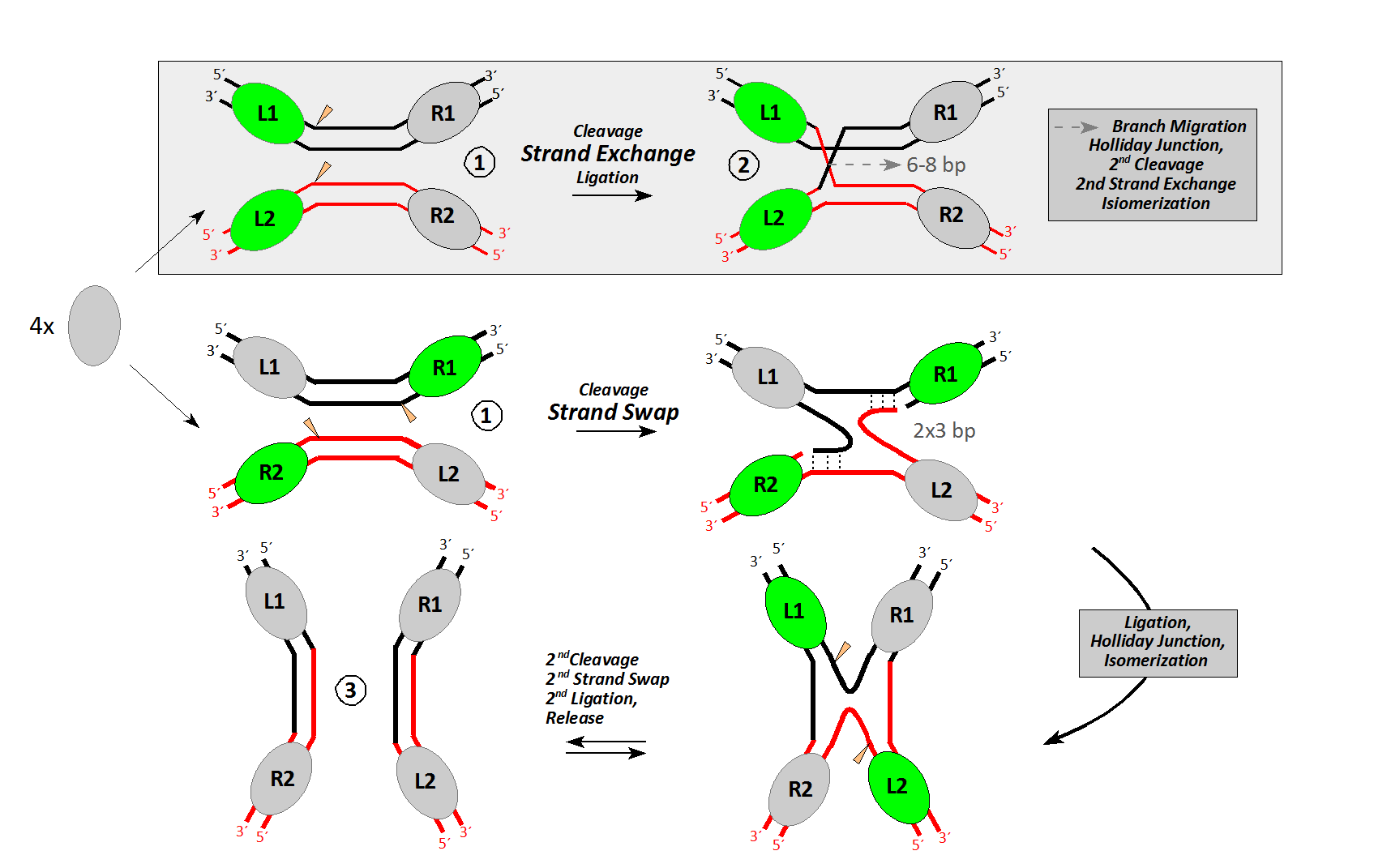
Setiap titik persilangan membentuk apa yang dikenal sebagai struktur Holliday (produk DNA salib dari peristiwa rekombinasi).
DNA salib ini harus diselesaikan oleh endonuklease lain. DNA chimeric atau hybrid dari struktur ini dapat diselesaikan dengan dua cara. Jika pemotongan endonukleotida kedua terjadi pada pita yang sama dengan yang pertama terjadi, tidak ada rekombinasi yang terjadi. Jika potongan kedua terjadi di pita lain, produk yang dihasilkan adalah rekombinan.
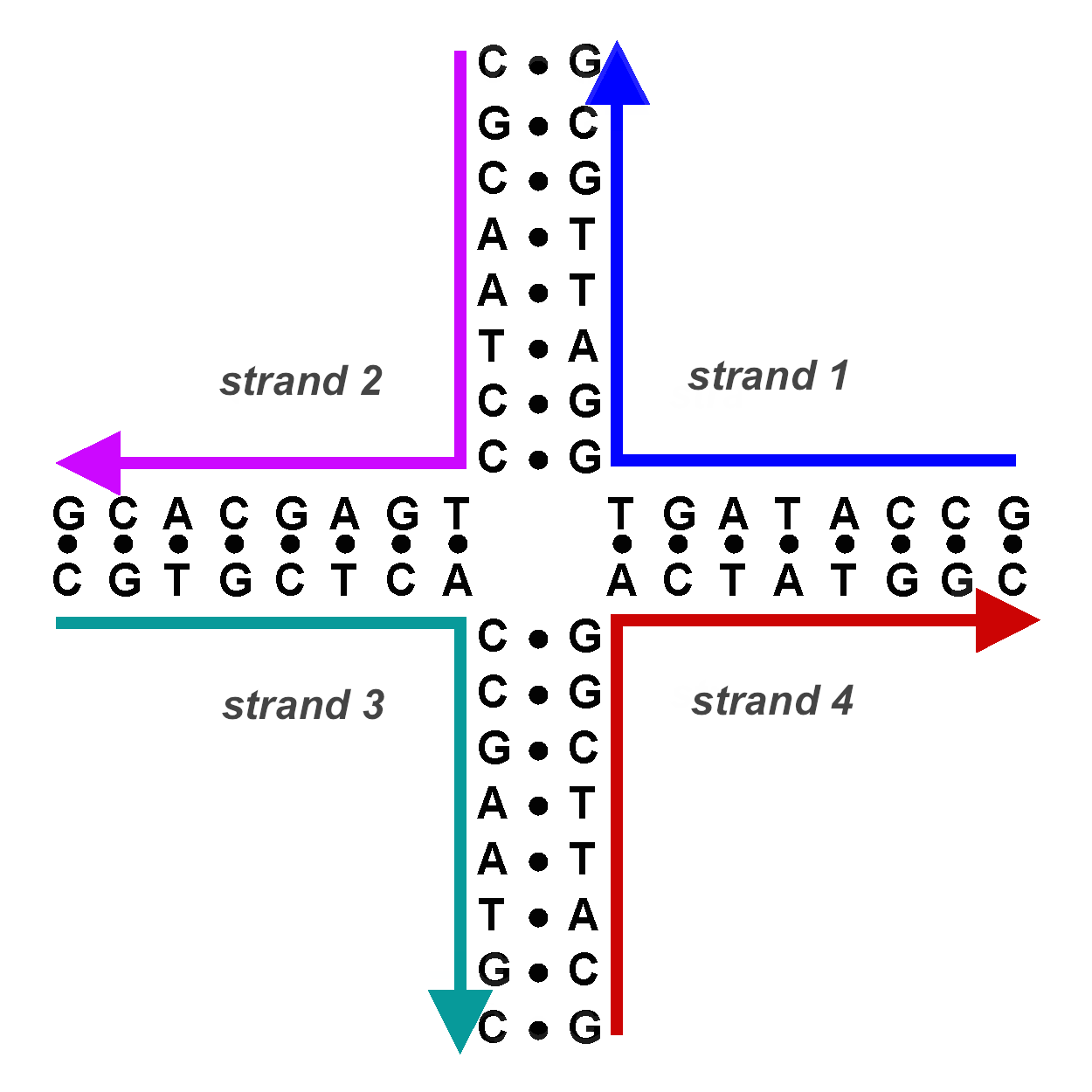
DNA rekombinan dalam struktur Holliday. es.m.wikipedia.org/wiki/File:Mao-4armjunction-schematic.png.
Rekombinasi V (D) J
Ini adalah jenis rekombinasi somatik (non-meiosis) yang berkontribusi pada pembentukan variabilitas antibodi sistem kekebalan yang sangat besar.
Rekombinasi ini terjadi pada fragmen tertentu dari gen yang mengkode rantai polipeptida yang mendefinisikannya. Ini dilakukan oleh sel B dan melibatkan daerah genetik yang berbeda.
Menariknya, ada parasit seperti Trypanosoma brucei yang menggunakan mekanisme rekombinasi serupa untuk menciptakan variabilitas antigen permukaan. Dengan cara ini, mereka dapat menghindari respons inang jika inang gagal menghasilkan antibodi yang mampu mengenali antigen “baru”.
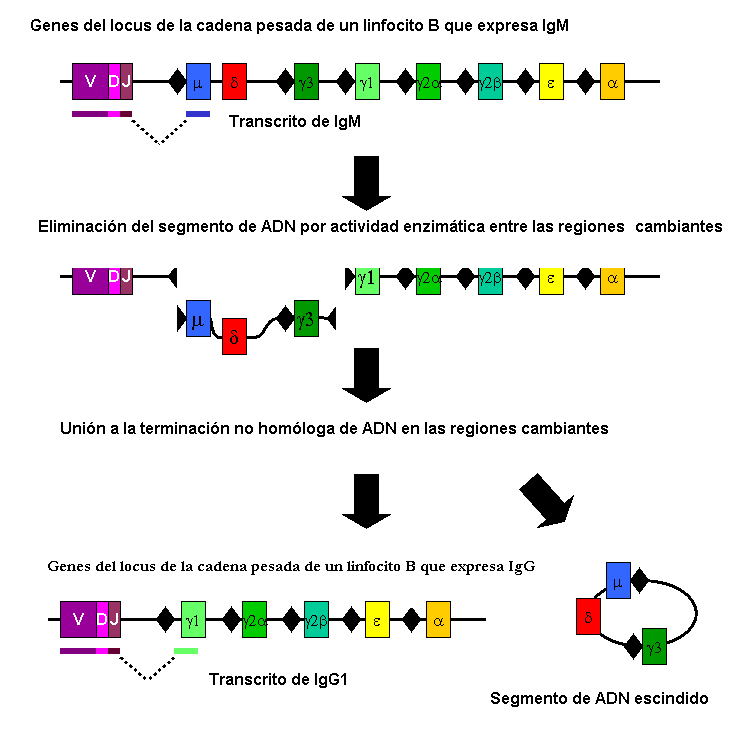
Keanekaragaman antibodi yang diciptakan oleh rekombinasi. es.m.wikipedia.org/wiki/File:Cambio_clase_recombinacion.PNG
-Rekombinasi non homolog
Akhirnya, ada proses rekombinasi yang tidak bergantung pada kesamaan urutan molekul yang berpartisipasi. Pada eukariota, misalnya, rekombinasi ujung non-homolog sangat penting.
Ini terjadi dengan fragmen DNA yang menunjukkan putusnya pita ganda pada DNA. Ini “diperbaiki” oleh sel yang menghubungkannya dengan fragmen lain juga dengan pemutusan pita ganda.
Namun, molekul-molekul ini tidak harus serupa untuk berpartisipasi dalam proses rekombinasi ini. Artinya, dengan memperbaiki kerusakan, sel dapat bergabung dengan DNA yang tidak terkait, sehingga menciptakan molekul (rekombinan) yang benar-benar baru.
Pentingnya rekombinasi
Pentingnya sebagai penyebab: replikasi dan perbaikan DNA
Rekombinasi memastikan ketepatan informasi DNA selama dan setelah proses replikasi. Rekombinasi mendeteksi kerusakan DNA selama proses pita baru dalam makromolekul yang sangat panjang ini.
Karena setiap pita memiliki informasinya sendiri, dan informasi pelengkapnya, rekombinasi menjamin bahwa tidak ada yang hilang. Masing-masing bertindak sebagai saksi bagi yang lain. Demikian pula, pada organisme diploid, kromosom homolog menyaksikan saudara kandungnya, dan sebaliknya.
Di sisi lain, sekali DNA telah direplikasi, mekanisme perbaikan kerusakan molekul ini bervariasi. Beberapa langsung (cedera ditindaklanjuti secara langsung) dan yang lain tidak langsung.
Mekanisme perbaikan tidak langsung bergantung pada rekombinasi yang terjadi. Artinya, untuk memperbaiki kerusakan pada molekul DNA, digunakan molekul homolog lain. Ini akan bertindak dalam rekombinasi reparatif sebagai templat dari mana ia telah mengalami kerusakan.
Pentingnya sebagai konsekuensi: generasi variabilitas genetik
Rekombinasi mampu menciptakan variabilitas kromosom yang sangat besar selama meiosis. Rekombinasi somatik juga menghasilkan variabilitas, seperti dalam kasus antibodi pada vertebrata .
Dalam banyak organisme meiosis adalah gametik. Dalam organisme yang bereproduksi secara seksual, rekombinasi ternyata menjadi salah satu cara paling ampuh untuk menghasilkan variabilitas.
Artinya, untuk mutasi spontan dan segregasi kromosom, rekombinasi harus ditambahkan sebagai unsur lain yang menghasilkan variabilitas gamet.
Integrasi genom bakteriofag dengan rekombinasi spesifik lokasi, di sisi lain, telah berkontribusi pada recaraling genom bakteri inang mereka.
Ini telah berkontribusi pada generasi variabilitas genom, dan evolusi, dari kelompok makhluk hidup yang penting ini.
Rekombinasi dan kesehatan
Kita telah melihat bahwa DNA dapat diperbaiki, tetapi bukan apa yang merusaknya. Pada kenyataannya, hampir semua hal dapat merusak DNA, dimulai dengan replikasi yang salah yang tidak diperbaiki.
Namun di luar itu, DNA dapat dirusak oleh sinar UV, radiasi pengion, radikal oksigen bebas yang dihasilkan oleh respirasi sel , dan apa yang kita makan, merokok, bernapas, menelan, atau menyentuh.
Untungnya, Anda tidak harus menyerah hidup untuk melindungi DNA. Anda harus melepaskan hal-hal tertentu, tetapi pekerjaan besar dilakukan oleh sel itu sendiri. Mekanisme untuk mendeteksi kerusakan DNA ini, dan perbaikannya, jelas memiliki dasar genetik, dan kekurangannya, memiliki konsekuensi yang sangat besar.
Penyakit yang berhubungan dengan cacat pada rekombinasi homolog termasuk, misalnya, sindrom Bloom dan Werner, kanker payudara dan ovarium familial, dll.
Referensi
- Alberts, B., Johnson, AD, Lewis, J., Morgan, D., Raff, M., Roberts, K., Walter, P. (2014) Biologi Molekuler Sel (Edisi ke-6). WW Norton & Company, New York, NY, AS.
- Bell, JC, Kowalczykowski, SC (2016) Mekanika dan interogasi molekul tunggal rekombinasi DNA. Tinjauan Tahunan Biokimia, 85: 193-226.
- Prado, F. () Rekombinasi Homolog: Untuk Fork dan Beyond. Gen, doi: 10.3390 / gen9120603
- Griffiths, AJF, Wessler, R., Carroll, SB, Doebley, J. (2015). Pengantar Analisis Genetika (Edisi ke-11). New York: WH Freeman, New York, NY, AS.
- Tock, AJ, Henderson, IR (2018) Hotspot untuk Inisiasi Rekombinasi Meiotik. Perbatasan dalam Genetika, doi: 10.3389 / fgene.2018.00521
- Wahl, A., Battesti, A., Ansaldi, M. (2018) Prophages in Salmonella enterica : kekuatan pendorong dalam membentuk kembali genom dan fisiologi inang bakteri mereka? Mikrobiologi Molekuler, doi: 10.1111 / mmi.14167.
- Wright, WD, Shah, SS, Heyer, WD (2018) Rekombinasi homolog dan perbaikan pemutusan untai ganda DNA. Jurnal Kimia Biologi, 293: 10524-10535
