
oksidasi beta dari asam lemak adalah rute dari katabolisme (degradasi) dari asam lemak, yang fungsi utamanya adalah produksi atau “release” dari energi yang terkandung dalam ikatan molekul-molekul ini.
Rute ini ditemukan pada tahun 1904 berkat percobaan yang dilakukan oleh Franz Knoop Jerman, yang terdiri dari pemberian asam lemak pada tikus percobaan yang gugus metil akhirnya telah dimodifikasi dengan gugus fenil.
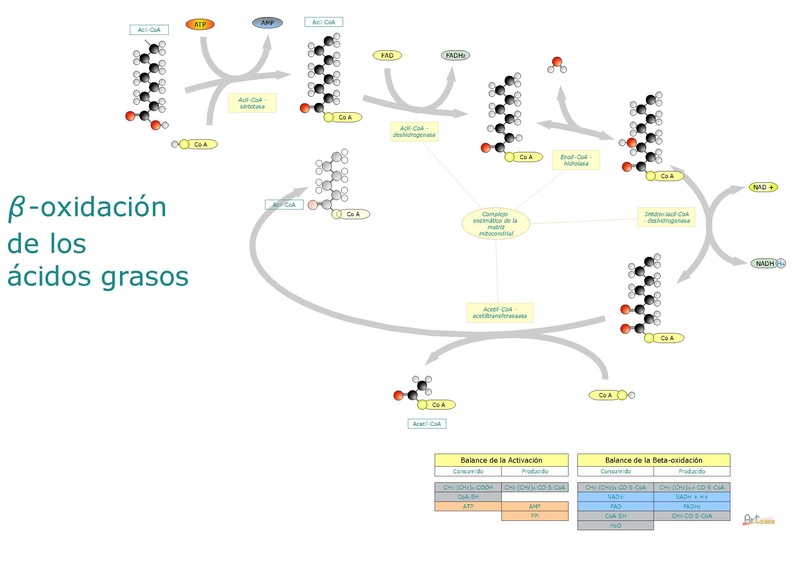
Diagram oksidasi beta asam lemak (Sumber: Arturo González Laguna [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)] melalui Wikimedia Commons)
Knoop mengharapkan produk katabolisme dari asam lemak “analog” ini mengikuti jalur yang mirip dengan jalur oksidasi asam lemak normal (alami yang tidak dimodifikasi). Namun, ia menemukan bahwa ada perbedaan dalam produk yang diperoleh sebagai fungsi dari jumlah atom karbon dari asam lemak.
Dengan hasil ini, Knoop mengusulkan bahwa degradasi terjadi dalam “langkah”, dimulai dengan “serangan” pada karbon (yang ada di posisi 3 sehubungan dengan gugus karboksil terminal), melepaskan fragmen dua atom karbon.
Kemudian ditunjukkan bahwa proses tersebut membutuhkan energi dalam bentuk ATP, yang diproduksi di mitokondria dan bahwa fragmen dua atom karbon memasuki siklus Krebs sebagai asetil-KoA.
Singkatnya, oksidasi beta asam lemak melibatkan aktivasi gugus karboksil terminal, pengangkutan asam lemak teraktivasi ke dalam matriks mitokondria, dan oksidasi dua-dua karbon dari gugus karboksil.
Seperti banyak proses anabolik dan katabolik, rute ini diatur, karena membutuhkan mobilisasi asam lemak “cadangan” ketika rute katabolik lainnya tidak cukup untuk memenuhi kebutuhan energi seluler dan tubuh.
Indeks artikel
Langkah dan reaksi
Asam lemak sebagian besar berada di sitosol, baik yang berasal dari jalur biosintetik atau dari timbunan lemak yang disimpan dari makanan yang dicerna (yang harus masuk ke dalam sel).
– Aktivasi asam lemak dan transportasi ke mitokondria
Aktivasi asam lemak memerlukan penggunaan molekul ATP dan berkaitan dengan pembentukan konjugat asil tioester dengan koenzim A.
Aktivasi ini dikatalisis oleh sekelompok enzim yang disebut ligase asetil-KoA khusus untuk panjang rantai masing-masing asam lemak. Beberapa dari enzim ini mengaktifkan asam lemak saat mereka diangkut ke dalam matriks mitokondria, karena mereka tertanam di membran luar mitokondria.
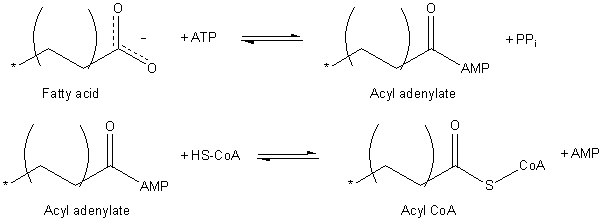
Aktivasi asam lemak (Sumber: Jag123 di Wikipedia bahasa Inggris [Domain publik] melalui Wikimedia Commons)
Proses aktivasi terjadi dalam dua langkah, pertama menghasilkan asil adenilat dari asam lemak teraktivasi dengan ATP, di mana molekul pirofosfat (PPi) dilepaskan. Gugus karboksil yang diaktifkan oleh ATP kemudian diserang oleh gugus tiol koenzim A untuk membentuk asil-KoA.
Translokasi asil-KoA melalui membran mitokondria dicapai berkat sistem transportasi yang dikenal sebagai antar-jemput karnitin.
– Oksidasi beta asam lemak jenuh dengan jumlah atom karbon genap
Degradasi asam lemak adalah rute siklus, karena pelepasan setiap fragmen dari dua atom karbon segera diikuti oleh yang lain, sampai panjang penuh molekul tercapai. Reaksi yang berperan dalam proses ini adalah sebagai berikut:
– Dehidrogenasi.
– Hidrasi ikatan rangkap.
– Dehidrogenasi gugus hidroksil.
– Fragmentasi dengan serangan molekul asetil-KoA pada karbon .
Reaksi 1: dehidrogenasi pertama
Ini terdiri dari pembentukan ikatan rangkap antara -karbon dan -karbon dengan menghilangkan dua atom hidrogen. Ini dikatalisis oleh enzim asil-KoA dehidrogenase, yang membentuk molekul trans∆2-enoyl-S-CoA dan molekul FAD + (kofaktor).
Reaksi 2 dan 3: hidrasi dan dehidrogenasi
Hidrasi dikatalisis oleh enoyl-CoA hydratase, sedangkan dehidrogenasi dimediasi oleh 3-hydroxyacyl-CoA dehydrogenase, dan reaksi terakhir tergantung pada kofaktor NAD +.
Hidrasi trans∆2-enoyl-S-CoA menghasilkan 3-hydroxyacyl-CoA, dehidrogenasinya menghasilkan molekul 3-ketoacyl-CoA dan NADH + H.
FADH2 dan NADH yang dihasilkan dalam tiga reaksi oksidasi beta pertama dioksidasi ulang melalui rantai transpor elektron, berkat mereka berpartisipasi dalam produksi ATP, 2 molekul untuk setiap FADH2 dan 3 molekul untuk setiap NADH.
Reaksi 4: fragmentasi
Setiap siklus oksidasi beta yang menghilangkan molekul dengan dua atom karbon berakhir dengan pembelahan “tiolitik” dari karbon keto, yang diserang oleh koenzim A pada ikatan antara karbon dan .
Reaksi ini dikatalisis oleh enzim -ketothiolase atau thiolase, dan produknya adalah satu molekul asil-KoA (asam lemak teraktivasi dengan dua atom karbon lebih sedikit) dan satu asetil-KoA.
– Oksidasi beta asam lemak jenuh dengan jumlah atom karbon ganjil
Pada asam lemak dengan jumlah atom karbon ganjil (yang jumlahnya tidak terlalu banyak), molekul pada siklus degradasi terakhir memiliki 5 atom karbon, sehingga fragmentasinya menghasilkan molekul asetil-KoA (yang memasuki siklus Krebs) dan molekul lain dari propionil-KoA.
Propionil-KoA harus dikarboksilasi (reaksi bergantung pada ATP dan bikarbonat) oleh enzim propionil-KoA karboksilase, sehingga membentuk senyawa yang dikenal sebagai D-metilmalonil-KoA, yang harus di-epimerisasi ke bentuk “L”.
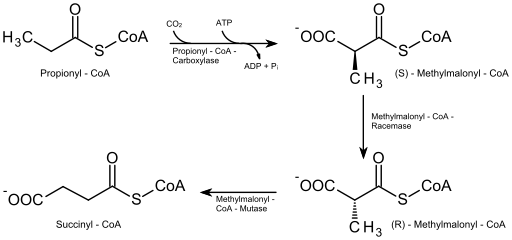
Oksidasi beta asam lemak bernomor ganjil (Sumber: Eleska [CC0] melalui Wikimedia Commons)
Senyawa yang dihasilkan dari epimerisasi kemudian diubah menjadi suksinil-KoA oleh aksi enzim L-metilmalonil-KoA mutase, dan molekul ini, serta asetil-KoA, memasuki siklus asam sitrat.
– Oksidasi beta asam lemak tak jenuh
Banyak lipid seluler memiliki rantai asam lemak tak jenuh, yaitu, mereka memiliki satu atau lebih ikatan rangkap di antara atom karbonnya.
Oksidasi asam lemak ini sedikit berbeda dengan asam lemak jenuh, karena dua enzim tambahan, enoyl-CoA isomerase dan 2,4-dienoyl-CoA reduktase, bertugas menghilangkan ketidakjenuhan ini sehingga asam lemak ini dapat substrat untuk enzim enoyl-CoA hydratase.
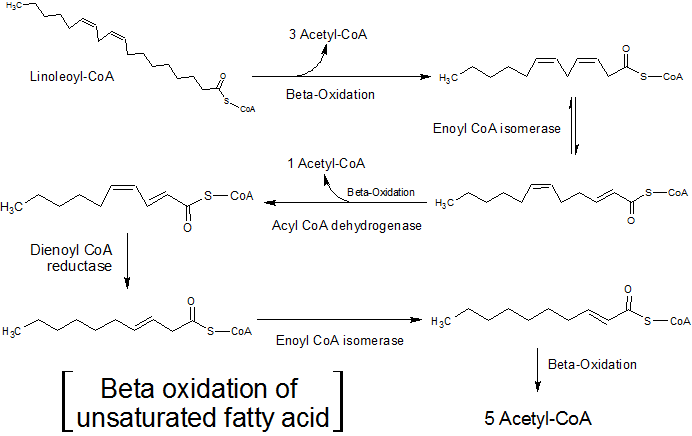
Oksidasi beta asam lemak tak jenuh (Sumber: Hajime7basketball [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)] melalui Wikimedia Commons)
Enoil-CoA isomerase bekerja pada asam lemak tak jenuh tunggal (dengan hanya satu ketidakjenuhan), sedangkan enzim 2,4-dienoil-KoA reduktase bereaksi dengan asam lemak tak jenuh ganda (dengan dua atau lebih ketidakjenuhan).
– Oksidasi ekstramitokondria beta
Oksidasi beta asam lemak juga dapat terjadi di dalam organel sitosol lain seperti peroksisom , misalnya, dengan perbedaan bahwa elektron yang ditransfer ke FAD + tidak dikirim ke rantai pernapasan, tetapi langsung ke oksigen.
Reaksi ini menghasilkan hidrogen peroksida (oksigen berkurang), suatu senyawa yang dihilangkan oleh enzim katalase, khusus untuk organel ini.
Produk oksidasi beta
Oksidasi asam lemak menghasilkan lebih banyak energi daripada pemecahan karbohidrat. Produk utama oksidasi beta adalah asetil-KoA yang dihasilkan di setiap langkah bagian siklik dari jalur tersebut, namun produk lainnya adalah:
– AMP, H + dan pirofosfat (PPi), diproduksi selama aktivasi.
– FADH2 dan NADH, untuk setiap asetil-KoA yang dihasilkan.
– Succinyl-CoA, ADP, Pi, untuk asam lemak rantai ganjil.
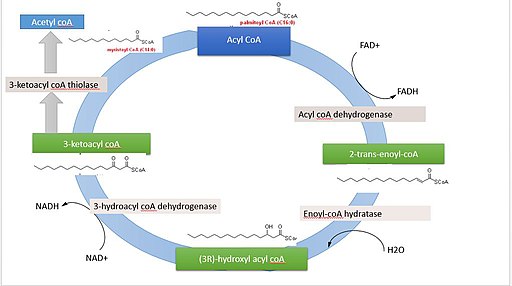
Oksidasi beta asam palmitat (Sumber: Rojinbkht [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)] melalui Wikimedia Commons)
Jika kita perhatikan sebagai contoh oksidasi beta lengkap asam palmitat (palmitat), asam lemak dengan 16 atom karbon, jumlah energi yang dihasilkan setara dengan kurang lebih 129 molekul ATP, yang berasal dari 7 putaran yang harus diselesaikan. .siklus.
Peraturan
Pengaturan oksidasi beta asam lemak di sebagian besar sel bergantung pada ketersediaan energi, tidak hanya terkait dengan karbohidrat tetapi juga dengan asam lemak itu sendiri.
Hewan mengontrol mobilisasi dan, oleh karena itu, pemecahan lemak melalui rangsangan hormonal, yang pada saat yang sama dikendalikan oleh molekul seperti cAMP, misalnya.
Di hati, organ pengurai lemak utama, konsentrasi malonil-KoA sangat penting untuk regulasi oksidasi beta; ini adalah substrat pertama yang dilakukan dalam jalur biosintesis asam lemak.
Ketika malonil-KoA terakumulasi dalam proporsi besar, itu mempromosikan biosintesis asam lemak dan menghambat transporter mitokondria atau antar-jemput asil-karnitin. Ketika konsentrasinya menurun, penghambatan berhenti dan oksidasi beta diaktifkan.
Referensi
- Mathews, C., van Holde, K., & Ahern, K. (2000). Biokimia (edisi ke-3). San Fransisco, California: Pearson.
- Nelson, DL, & Cox, MM (2009). Prinsip Biokimia Lehninger . Edisi Omega ( edisi ke-5).
- Rawn, JD (1998). Biokimia . Burlington, Massachusetts: Penerbit Neil Patterson.
- Schulz, H. (1991). Oksidasi beta asam lemak Biochimica et Biophysica Acta , 1081 , 109–120.
- Schulz, H. (1994). Pengaturan Oksidasi Asam Lemak di Jantung. Tinjauan Kritis , 165-171.
- Schulz, H., & Kunau, W. (1987). Beta-oksidasi asam lemak tak jenuh: jalur direvisi. TIBS , 403–406.
