
fosforilasi oksidatif adalah proses di mana molekul disintesis ATP dari ADP dan P i (fosfat anorganik). Mekanisme ini dilakukan oleh bakteri dan sel eukariotik . Dalam sel eukariotik, fosforilasi terjadi di matriks mitokondria sel non-fotosintetik.
Produksi ATP didorong oleh transfer elektron dari koenzim NADH atau FADH 2 ke O 2 . Proses ini merupakan produksi energi tertinggi dalam sel dan berasal dari pemecahan karbohidrat dan lemak.
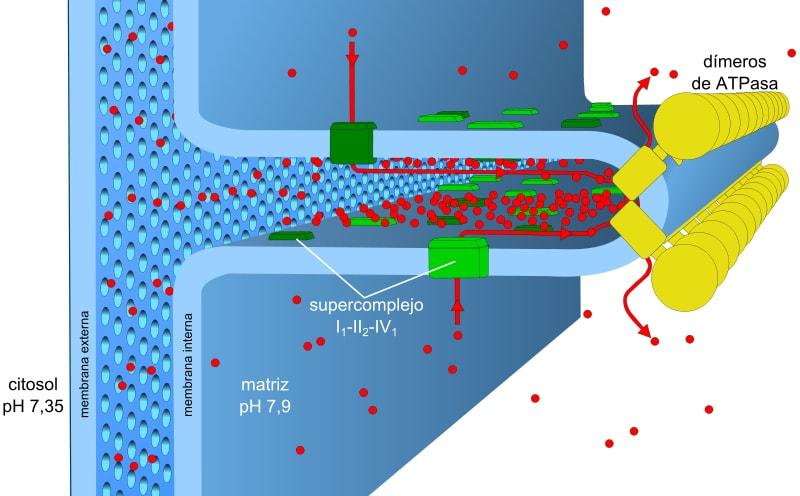
Sumber: Robot8A [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)]
Energi yang tersimpan dalam muatan dan gradien pH, juga dikenal sebagai gaya gerak proton, memungkinkan proses ini berlangsung. Gradien proton yang dihasilkan menyebabkan bagian luar membran bermuatan positif karena konsentrasi proton (H + ) dan matriks mitokondria menjadi negatif.
Indeks artikel
Di mana fosforilasi oksidatif terjadi?
Proses transpor elektron dan fosforilasi oksidatif berhubungan dengan membran. Pada prokariota mekanisme ini dilakukan melalui membran plasma . Dalam sel eukariotik mereka berhubungan dengan membran mitokondria .
Jumlah mitokondria yang ditemukan dalam sel bervariasi tergantung pada jenis sel. Misalnya, pada mamalia , eritrosit kekurangan organel ini, sedangkan jenis sel lain, seperti sel otot, bisa mencapai jutaan.
Membran mitokondria terdiri dari membran luar yang sederhana, membran dalam yang agak lebih kompleks, dan di antara keduanya terdapat ruang antar membran, tempat banyak enzim yang bergantung pada ATP berada.
Membran luar mengandung protein yang disebut porin yang membentuk saluran untuk difusi sederhana molekul kecil. Membran ini bertanggung jawab untuk menjaga struktur dan bentuk mitokondria.
Membran bagian dalam memiliki kepadatan yang lebih tinggi dan kaya akan protein. Ini juga kedap untuk molekul dan ion sehingga, untuk melewatinya, mereka membutuhkan protein antar membran untuk mengangkutnya.
Di dalam matriks, lipatan membran bagian dalam memanjang, membentuk punggungan yang memungkinkannya memiliki area yang luas dalam volume yang kecil .
Pembangkit listrik sel
Mitokondria dianggap sebagai pusat penghasil energi seluler. Ini berisi enzim yang terlibat dalam proses siklus asam sitrat, oksidasi asam lemak, dan enzim redoks dan protein untuk transpor elektron dan fosforilasi ADP.
Gradien konsentrasi proton (gradien pH) dan gradien muatan atau potensial listrik di membran dalam mitokondria bertanggung jawab atas gaya gerak proton. Permeabilitas membran dalam yang rendah untuk ion (selain H + ) memungkinkan mitokondria memiliki gradien tegangan yang stabil.
Transpor elektronik, pemompaan proton, dan perolehan ATP terjadi secara bersamaan di mitokondria, berkat gaya gerak proton. Gradien pH mempertahankan kondisi asam di intermembran dan di matriks mitokondria dengan kondisi basa.
Untuk setiap dua elektron dipindahkan ke O 2 sekitar 10 proton dipompa melalui membran, menciptakan gradien elektrokimia. Energi yang dilepaskan dalam proses ini dihasilkan secara bertahap oleh perjalanan elektron melalui rantai transpor.
Tahapan
Energi yang dilepaskan selama reaksi oksidasi-reduksi dari NADH dan FADH 2 adalah cukup tinggi (sekitar 53 kkal / mol untuk setiap pasangan elektron), sehingga untuk digunakan dalam pembuatan molekul ATP, harus diproduksi secara bertahap dengan berlalunya elektron melalui transporter.
Ini diatur menjadi empat kompleks yang terletak di membran mitokondria bagian dalam. Penggabungan reaksi-reaksi ini dengan sintesis ATP terjadi di kompleks kelima.
Rantai transpor elektron
NADH mentransfer sepasang elektron yang memasuki kompleks I dari rantai transpor elektron. Elektron ditransfer ke flavin mononukleotida, dan kemudian ke ubiquinone (koenzim Q) melalui transporter besi-sulfur. Proses ini melepaskan sejumlah besar energi (16,6 kkal/mol).
Ubiquinone mengangkut elektron melintasi membran ke kompleks III. Dalam kompleks ini, elektron melewati sitokrom b dan c 1 berkat transporter besi-sulfur.
Elektron berpindah dari kompleks III ke kompleks IV (sitokrom c oksidase), ditransfer satu per satu dalam sitokrom c (protein membran perifer). Dalam kompleks IV elektron melewati sepasang ion tembaga (Cu a 2+ ), kemudian ke sitokrom c a , lalu ke pasangan ion tembaga lainnya (Cu b 2+ ) dan dari sini ke sitokrom a 3 .
Akhirnya, elektron ditransfer ke O 2 yang merupakan akseptor terakhir dan membentuk molekul air (H 2 O) untuk setiap pasangan elektron yang diterima. Bagian elektron dari IV kompleks untuk O 2 juga menghasilkan sejumlah besar energi bebas (25,8 kkal / mol).
Suksinat CoQ reduktase
Kompleks II (suksinat CoQ reduktase) menerima pasangan elektron dari siklus asam sitrat, melalui oksidasi molekul suksinat menjadi fumarat. Elektron ini ditransfer ke FAD, kemudian melalui kelompok besi-sulfur, ke ubiquinone. Dari koenzim ini mereka melewati kompleks III dan mengikuti rute yang dijelaskan sebelumnya.
Energi yang dilepaskan dalam reaksi transfer elektron ke FAD tidak cukup untuk mendorong proton melalui membran, sehingga tidak ada gaya gerak proton yang dihasilkan pada langkah rantai ini, dan akibatnya FADH menghasilkan lebih sedikit H + daripada NADH.
Kopling atau transduksi energi
Energi yang dihasilkan dalam proses transpor elektron yang dijelaskan sebelumnya harus dapat digunakan untuk produksi ATP, suatu reaksi yang dikatalisis oleh enzim ATP sintase atau kompleks V. Kekekalan energi ini dikenal sebagai energy coupling, dan mekanismenya telah sulit untuk dikarakterisasi.
Beberapa hipotesis telah dijelaskan untuk menggambarkan transduksi energi ini. Yang terbaik diterima adalah hipotesis kopling kemosmotik, dijelaskan di bawah ini.
Kopling kemosmotik
Mekanisme ini mengusulkan bahwa energi yang digunakan untuk sintesis ATP berasal dari gradien proton dalam membran sel. Proses ini campur tangan dalam mitokondria, kloroplas dan bakteri dan terkait dengan pengangkutan elektron.
Kompleks I dan IV dari transpor elektron bertindak sebagai pompa proton. Ini mengalami perubahan konformasi yang memungkinkan mereka untuk memompa proton ke dalam ruang antarmembran. Dalam IV kompleks, untuk setiap pasangan elektron, dua proton dipompa keluar dari membran dan dua lagi tetap dalam matriks, membentuk H 2 O.
Ubiquinone di kompleks III menerima proton dari kompleks I dan II dan melepaskannya ke luar membran. Kompleks I dan III masing-masing memungkinkan lewatnya empat proton untuk setiap pasangan elektron yang diangkut.
Matriks mitokondria memiliki konsentrasi proton yang rendah dan potensial listrik negatif, sedangkan ruang antarmembran menyajikan kondisi terbalik. Aliran proton melalui membran ini mewakili gradien elektrokimia yang menyimpan energi yang diperlukan (± 5 kkal / mol per proton) untuk sintesis ATP.
Sintesis ATP
Enzim ATP sintetase adalah kompleks kelima yang terlibat dalam fosforilasi oksidatif. Ini bertanggung jawab untuk memanfaatkan energi gradien elektrokimia untuk membentuk ATP.
Protein transmembran ini terdiri dari dua komponen: F 0 dan F 1 . Komponen F 0 memungkinkan kembalinya proton ke matriks mitokondria, berfungsi sebagai saluran dan F 1 mengkatalisis sintesis ATP melalui ADP dan P i , menggunakan energi pengembalian tersebut.
Proses sintesis ATP membutuhkan perubahan struktural pada F 1 dan perakitan komponen F 0 dan F 1 . Translokasi proton melalui F 0 menyebabkan perubahan konformasi pada tiga subunit F 1 , memungkinkannya bertindak sebagai motor rotasi, mengarahkan pembentukan ATP.
Subunit yang bertanggung jawab untuk pengikatan ADP dengan P i beralih dari keadaan lemah (L) ke keadaan aktif (T). Ketika ATP terbentuk, subunit kedua masuk ke keadaan terbuka (O) yang memungkinkan pelepasan molekul ini. Setelah ATP dilepaskan, subunit ini beralih dari keadaan terbuka ke keadaan tidak aktif (L).
Molekul ADP dan P i mengikat subunit yang telah berpindah dari keadaan O ke keadaan L.
Produk
Rantai transpor elektron dan fosforilasi menghasilkan molekul ATP. Oksidasi NADH menghasilkan sekitar 52,12 kkal / mol (218 kJ / mol) energi bebas.
Reaksi keseluruhan untuk oksidasi NADH adalah:
NADH + 1/2 O 2 + H + ↔ H 2 O + NAD +
Transfer elektron dari NADH dan FADH 2 terjadi melalui berbagai kompleks, memungkinkan perubahan energi bebas G ° untuk dipecah menjadi “paket” energi yang lebih kecil yang digabungkan ke sintesis ATP.
Oksidasi satu molekul NADH menghasilkan sintesis tiga molekul ATP. Sedangkan oksidasi dari FADH 2 molekul digabungkan dengan sintesis dua ATP.
Koenzim ini berasal dari proses glikolisis dan siklus asam sitrat. Untuk setiap molekul glukosa yang terdegradasi, mereka akhirnya menghasilkan 36 atau 38 molekul ATP, tergantung pada lokasi sel. Di otak dan otot rangka 36 ATP diproduksi sedangkan di jaringan otot 38 ATP diproduksi.
Fitur
Semua organisme, uniseluler dan multiseluler, membutuhkan energi minimal dalam sel mereka untuk melakukan proses di dalamnya, dan pada gilirannya mempertahankan fungsi vital di seluruh organisme.
Proses metabolisme membutuhkan energi untuk berlangsung. Sebagian besar energi yang dapat digunakan diperoleh dari pemecahan karbohidrat dan lemak. Energi ini berasal dari proses fosforilasi oksidatif.
Kontrol fosforilasi oksidatif
Tingkat pemanfaatan ATP dalam sel mengontrol sintesis, dan pada gilirannya, karena kopling fosforilasi oksidatif dengan rantai transpor elektron, juga umumnya mengatur laju transpor elektron.
Fosforilasi oksidatif memiliki kontrol ketat yang memastikan bahwa ATP tidak dihasilkan lebih cepat daripada yang dikonsumsi. Ada langkah-langkah tertentu dalam proses transpor elektron dan fosforilasi berpasangan yang mengatur laju produksi energi.
Kontrol terkoordinasi dari produksi ATP
Jalur utama produksi energi (ATP seluler) adalah glikolisis , siklus asam sitrat, dan fosforilasi oksidatif. Kontrol terkoordinasi dari ketiga proses ini mengatur sintesis ATP.
Kontrol fosforilasi oleh rasio aksi massa ATP tergantung pada pasokan elektron yang tepat dalam rantai transpor. Ini pada gilirannya tergantung pada rasio [NADH] / [NAD + ] , yang dijaga tetap tinggi oleh aksi glikolisis dan siklus asam sitrat.
Kontrol terkoordinasi ini dilakukan dengan mengatur titik kontrol glikolisis (PFK dihambat oleh sitrat) dan siklus asam sitrat (piruvat dehidrogenase, sitrat tapease, isositrat dehidrogenase dan -ketoglutarat dehidrogenase).
Kontrol oleh akseptor
Kompleks IV (sitokrom c oksidase) adalah enzim yang diatur oleh salah satu substratnya, yaitu aktivitasnya dikendalikan oleh sitokrom c tereduksi (c 2+ ), yang pada gilirannya berada dalam kesetimbangan dengan rasio konsentrasi antara [NADH] / [ Rasio NAD + ] dan aksi massa [ATP] / [ADP] + [P i ].
Semakin tinggi rasio [NADH] / [NAD + ] dan semakin rendah [ATP] / [ADP] + [P i ], semakin banyak konsentrasi sitokrom [c 2+ ] dan semakin besar aktivitas kompleks IV. Ini ditafsirkan, misalnya, jika kita membandingkan organisme dengan aktivitas istirahat dan aktivitas tinggi yang berbeda.
Pada individu dengan aktivitas fisik yang tinggi, konsumsi ATP dan oleh karena itu hidrolisisnya menjadi ADP + P i akan sangat tinggi, menghasilkan perbedaan rasio aksi massa yang menyebabkan peningkatan [c 2+ ] dan oleh karena itu keduanya meningkatkan sintesis ATP. Pada individu saat istirahat, situasi sebaliknya terjadi.
Pada akhirnya, tingkat fosforilasi oksidatif meningkat dengan konsentrasi ADP dalam mitokondria. Mengatakan konsentrasi tergantung pada translocators ADP-ATP bertanggung jawab untuk pengangkutan nukleotida adenin dan P i dari sitosol ke matriks mitokondria.
Agen pemutus
Fosforilasi oksidatif dipengaruhi oleh bahan kimia tertentu, yang memungkinkan transpor elektron berlanjut tanpa terjadi fosforilasi ADP, melepaskan produksi dan konservasi energi.
Agen-agen ini merangsang tingkat konsumsi oksigen mitokondria tanpa adanya ADP, juga menyebabkan peningkatan hidrolisis ATP. Mereka bekerja dengan menghilangkan perantara atau memutus status energi dalam rantai transpor elektron.
2,4-dinitrofenol, asam lemah yang melewati membran mitokondria, bertanggung jawab untuk menghilangkan gradien proton, karena mengikat mereka di sisi asam dan melepaskannya di sisi basa.
Senyawa ini digunakan sebagai “pil diet” karena ditemukan menghasilkan peningkatan respirasi, oleh karena itu peningkatan laju metabolisme dan penurunan berat badan yang terkait . Namun, ternyata efek negatifnya malah bisa menyebabkan kematian.
Disipasi gradien proton menghasilkan panas. Sel-sel dalam jaringan adiposa coklat menggunakan pelepasan yang dikontrol secara hormonal untuk menghasilkan panas. Mamalia berhibernasi dan bayi baru lahir yang tidak memiliki rambut terdiri dari jaringan ini yang berfungsi sebagai semacam selimut termal.
Inhibitor
Senyawa atau agen penghambat mencegah kedua konsumsi O 2 (elektron transportasi) dan fosforilasi oksidatif yang terkait. Agen ini mencegah pembentukan ATP dengan menggunakan energi yang dihasilkan dalam transportasi elektronik. Oleh karena itu, rantai transportasi berhenti ketika konsumsi energi tersebut tidak tersedia.
Antibiotik oligomisin berfungsi sebagai penghambat fosforilasi pada banyak bakteri, mencegah stimulasi ADP menjadi sintesis ATP.
Ada juga agen ionofor, yang membentuk kompleks yang larut dalam lemak dengan kation seperti K + dan Na + , dan melewati membran mitokondria dengan kation ini. Mitokondria kemudian menggunakan energi yang dihasilkan dalam transpor elektron untuk memompa kation alih-alih mensintesis ATP.
Referensi
- Alberts, B., Bray, D., Hopkin, K., Johnson, A., Lewis, J., Raff, M., Roberts, K. & Walter, P. (2004). Biologi sel esensial . New York: Ilmu Garland.
- Cooper, GM, Hausman, RE & Wright, N. (2010). Sel. (hal. 397-402). Marban.
- Devlin, TM (1992). Buku teks biokimia: dengan korelasi klinis. John Wiley & Sons, Inc.
- Garrett, RH, & Grisham, CM (2008). Biokimia. Thomson Brooks / Cole.
- Lodish, H., Darnell, JE, Berk, A., Kaiser, CA, Krieger, M., Scott, MP, & Matsudaira, P. (2008). Biologi sel molekuler. Macmillan.
- Nelson, DL, & Cox, MM (2006). Prinsip-prinsip Biokimia Lehninger edisi ke-4 . Ed Omega. Barcelona.
- Voet, D., & Voet, JG (2006). Biokimia. Ed. Medis Panamerika.
