
kromatin adalah kompleks yang dibentuk oleh DNA dan protein hanya dalam organisme eukariotik. Dalam hal proporsi, mengandung hampir dua kali lebih banyak protein daripada materi genetik. Protein yang paling penting dalam kompleks ini adalah histon – kecil, protein bermuatan positif yang mengikat DNA melalui interaksi elektrostatik. Selain itu, kromatin memiliki lebih dari seribu protein selain histon.
Unit dasar kromatin adalah nukleosom, yang terdiri dari penyatuan histon dan DNA. Susunan ini mengingatkan pada manik-manik pada kalung. Setelah melalui semua tingkat organisasi DNA yang lebih tinggi, kita akan sampai pada kromosom.
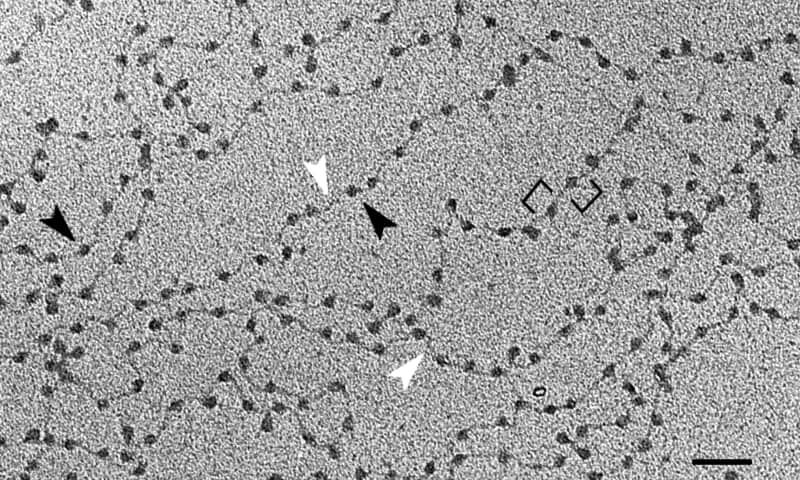
Sumber: Chromatin_nucleofilaments.png: Chris Woodcock Karya turunan: Gouttegd [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)]
Struktur kromatin berkaitan erat dengan kontrol ekspresi gen. Ada dua jenis utama: eukromatin dan heterokromatin.
Eukromatin dicirikan dengan memiliki tingkat pemadatan yang rendah, yang diterjemahkan ke dalam tingkat transkripsi yang tinggi. Sebaliknya, heterokromatin tidak aktif secara transkripsi, karena tingkat pemadatannya yang tinggi.
Secara struktural ada tanda epigenetik tertentu dari histon yang khas dari kedua jenis kromatin. Sementara eukromatin dikaitkan dengan asetilasi, heterokromatin dikaitkan dengan penurunan gugus kimia.
Ada beberapa daerah heterokromatin dengan fungsi struktural, seperti telomer dan sentromer.
Indeks artikel
Perspektif sejarah
Studi materi genetik dan organisasi strukturalnya dimulai pada tahun 1831 ketika peneliti Robert Brown mendeskripsikan nukleus. Salah satu pertanyaan langsung untuk penemuan ini adalah untuk menanyakan tentang sifat biologis dan kimia dari struktur ini.
Pertanyaan-pertanyaan ini mulai dijelaskan antara tahun 1870 dan 1900, dengan eksperimen Friedrich Miescher, yang memperkenalkan kata nuklein. Namun, Walther Flemming memodifikasi istilah tersebut dan menggunakan kromatin untuk merujuk pada zat inti.
Dengan berlalunya waktu, seseorang mulai memiliki pengetahuan yang lebih dalam tentang materi genetik dan sifat-sifatnya. Baru pada tahun 1908 ketika peneliti Italia Pasquale Baccarini memperhatikan bahwa kromatin tidak homogen dan berhasil memvisualisasikan benda-benda kecil di dalam nukleus.
Jenis kromatin – eukromatin dan heterokromatin – awalnya diusulkan oleh Emil Heitz pada tahun 1928. Untuk menetapkan klasifikasi ini, Heitz mengandalkan penggunaan pewarna.
Pada tahun 1974 ahli biologi Roger Kornberg mengusulkan caral untuk organisasi materi genetik dalam struktur yang dikenal sebagai nukleosom, hipotesis dikonfirmasi secara empiris oleh eksperimen Markus Noll.
Apa itu kromatin?
Komponen kromatin: DNA dan protein
Kromatin adalah nukleoprotein yang dibentuk oleh penyatuan materi genetik – DNA – dengan seperangkat protein yang heterogen. Asosiasi ini sangat dinamis dan memperoleh konformasi tiga dimensi yang kompleks yang memungkinkannya menjalankan fungsi regulasi dan strukturalnya.
Salah satu protein terpenting dalam kromatin adalah histon, yang ditemukan dalam proporsi yang hampir sama dengan DNA.
Histon adalah protein dasar, sangat dilestarikan sepanjang sejarah evolusi makhluk organik – yaitu, histon kita tidak jauh berbeda dibandingkan dengan mamalia lain, bahkan hewan lain yang lebih jauh secara filogenetik.
Muatan histon adalah positif, sehingga mereka dapat berinteraksi melalui gaya elektrostatik dengan muatan negatif dari tulang punggung fosfat yang ada dalam DNA. Ada lima jenis histon, yaitu: H1, H2A, H2B, H3, dan H4.
Ada juga sejumlah protein yang sifatnya berbeda dari histon yang berpartisipasi dalam pemadatan DNA.
Pemadatan DNA: nukleosom
Unit dasar kromatin adalah nukleosom – struktur berulang yang terdiri dari DNA dan histon, suatu konformasi yang ditemukan di seluruh materi genetik.
Heliks ganda DNA dilukai menjadi kompleks delapan histon yang dikenal sebagai oktamer histon . Molekul melilit sekitar dua putaran, diikuti oleh daerah pendek (antara 20 dan 60 pasangan basa) yang memisahkan nukleosom satu sama lain.
Untuk memahami organisasi ini, kita harus memperhitungkan bahwa molekul DNA sangat panjang (sekitar 2 meter) dan harus dililitkan secara teratur untuk menempatkan dirinya di dalam nukleus (yang diameternya 3 hingga 10 m). Selanjutnya, itu harus tersedia untuk replikasi dan transkripsi.
Tujuan ini dicapai dengan berbagai tingkat pemadatan DNA, yang pertama adalah nukleosom yang disebutkan di atas. Ini menyerupai manik-manik pada kalung mutiara. Kira-kira 150 pasangan basa DNA dililitkan di “jumlah” rumah.
Tidak ada kisah nyata dalam bakteri. Sebaliknya, ada serangkaian protein yang mengingatkan pada histon dan ini dianggap berkontribusi pada pengemasan DNA bakteri.
Organisasi yang unggul
Organisasi kromatin tidak dibatasi pada tingkat nukleosom. Asosiasi protein dan DNA ini dikelompokkan bersama dalam struktur yang lebih tebal sekitar 30 nm – karena ketebalan ini disebut tingkat “serat 30 nm”.
Kromatin yang diatur dalam ketebalan 30 nm diatur secara bergantian dalam bentuk loop yang memanjang dalam semacam perancah yang bersifat protein (bukan histon).
Ini adalah caral yang saat ini digunakan, meskipun keberadaan mekanisme pemadatan yang lebih kompleks dapat diharapkan. Organisasi terakhir terdiri dari kromosom.
Kesalahan dalam organisasi kromatin
Pemadatan dan pengorganisasian materi genetik sangat penting untuk berbagai fungsi biologis. Berbagai kondisi medis telah dikaitkan dengan kesalahan dalam struktur kromatin, di antaranya talasemia alfa terkait-X, sindrom Rubinstein-Taybi, sindrom Coffin-Lowry, sindrom Rett, dan lain-lain.
Jenis kromatin
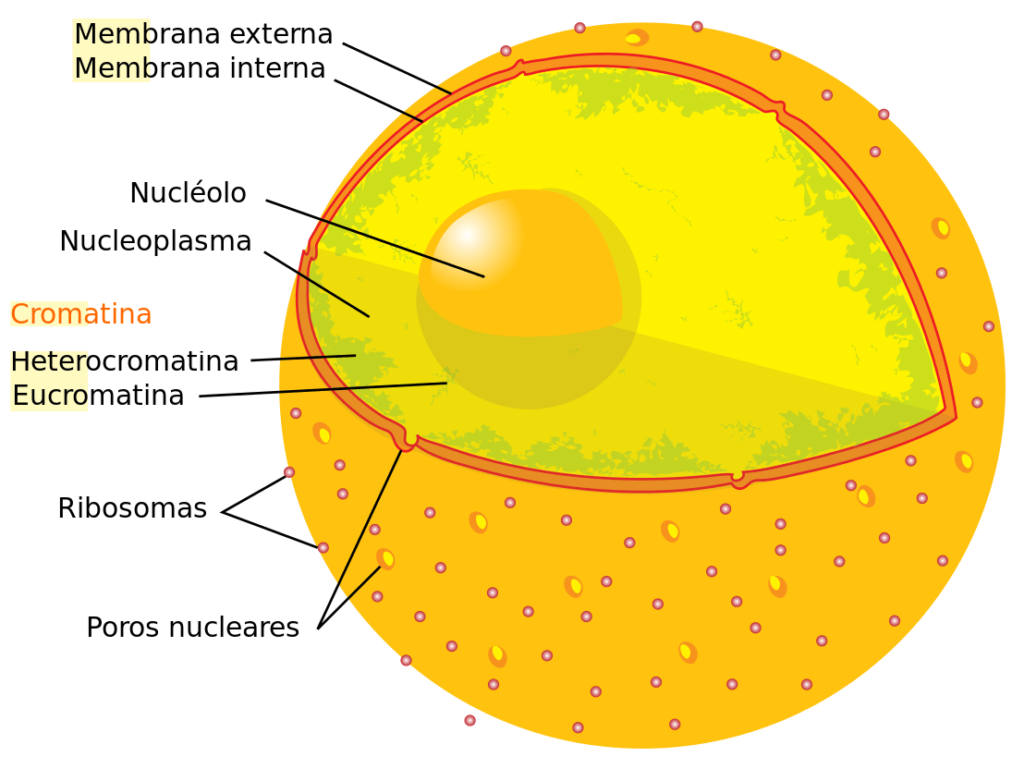
Ada dua jenis kromatin di dalam sel, yang terlihat dari aplikasi pewarnaan: eukromatin (kromatin “benar”) dan heterokromatin. Dalam kasus pertama, pewarnaannya diamati dengan lemah, sedangkan pada kasus kedua pewarnaannya intens.
Organisasi struktural DNA ini unik untuk organisme eukariotik dan sangat penting untuk perilaku kromosom dan regulasi ekspresi gen.
Jika kita mengevaluasi proporsi kedua jenis kromatin dalam sel yang berada di interfase, kita menemukan bahwa sekitar 90% dari kromatin adalah eukromatin dan 10% sisanya sesuai dengan heterokromatin. Kita akan menjelaskan setiap jenis secara rinci di bawah ini:
I. Heterokromatin
Karakteristik
Perbedaan utama antara kedua jenis kromatin terkait dengan tingkat pemadatan atau “pengemasan” molekul selama tahap tertentu dalam pembelahan sel.
Meskipun pada antarmuka materi genetik tampak tersebar secara acak, tidak demikian.
Ada organisasi yang signifikan pada tahap ini, di mana partisi diferensial dari bahan kromosom dapat dilihat di dalam nukleus.
DNA kromosom tidak terjalin dengan untai DNA kromosom lain dan tetap berada di daerah tertentu yang disebut wilayah kromosom. Organisasi ini tampaknya berkontribusi pada ekspresi gen.
Heterochromatin sangat kental, sehingga tidak dapat diakses oleh mesin transkripsi – sehingga tidak ditranskripsi. Juga, miskin dalam hal jumlah gen yang dimilikinya.
Jenis-jenis heterokromatin
Daerah tertentu dari heterokromatin bersifat persisten di sepanjang garis keturunan sel – yaitu, mereka akan selalu berperilaku seperti heterokromatin. Jenis heterokromatin ini dikenal sebagai konstitutif. Contohnya adalah daerah padat kromosom yang disebut sentromer dan telomer.
Sebaliknya, ada bagian heterokromatin yang dapat memvariasikan tingkat pemadatan sebagai respons terhadap perubahan pola perkembangan atau variabel lingkungan.
Berkat studi baru, pandangan ini sedang dirumuskan kembali dan sekarang ada bukti bahwa heterokromatin konstitutif juga dinamis dan mampu merespons rangsangan.
Struktur
Salah satu faktor yang menentukan struktur kromatin adalah modifikasi kimia histon. Dalam kasus kromatin yang tidak aktif secara transkripsi, mereka menunjukkan histon hipoasetilasi.
Penurunan jumlah gugus asetil dikaitkan dengan pembungkaman gen, karena tidak menutupi muatan positif lisin, memungkinkan interaksi elektrostatik yang kuat antara DNA dan histon.
Merek epigenetik lainnya adalah metilasi. Namun, karena penambahan gugus metil tidak mengubah muatan protein, konsekuensinya (gen pengaktifan atau penonaktifan) tidak begitu jelas dan akan tergantung pada daerah histon tempat tanda ditemukan.
Secara empiris telah ditemukan bahwa metilasi H3K4me3 dan H3K36me3 dikaitkan dengan aktivasi gen dan metilasi H3K9me3 dan H3K27me3.
Fitur
Dalam contoh heterokromatin konstitutif kita menyebutkan sentromer. Wilayah kromosom ini memiliki peran struktural dan berkontribusi pada pergerakan kromosom selama peristiwa pembelahan sel mitosis dan meiosis.
II. Eukromatin
Karakteristik
Berlawanan dengan heterokromatin, eukromatin adalah molekul yang kurang kompak, sehingga mesin transkripsi memiliki akses yang mudah (khususnya enzim RNA polimerase) dan dapat diekspresikan secara aktif melalui jalur genetik.
Struktur
Struktur nukleosom dari kromatin aktif transkripsi dicirikan oleh histon asetat. Kehadiran lisin monometilasi juga dikaitkan dengan aktivasi gen.
Penambahan gugus asetil ke residu histon lisin ini menetralkan muatan positif asam amino tersebut. Konsekuensi langsung dari perubahan ini adalah pengurangan interaksi elektrostatik antara histon dan DNA, menghasilkan kromatin yang lebih longgar.
Modifikasi struktural ini memungkinkan interaksi materi genetik dengan mesin transkripsi, yang ditandai dengan menjadi sangat besar.
Fitur
Eukromatin mencakup semua gen yang aktif dan berhasil dicapai oleh mesin enzimatik yang terkait dengan transkripsi. Oleh karena itu, fungsinya sama luasnya dengan fungsi gen yang terlibat.
Referensi
- Grewal, SI, & Moazed, D. (2003). Heterochromatin dan kontrol epigenetik ekspresi gen. sains , 301 (5634), 798-802.
- Jost, KL, Bertulat, B., & Cardoso, MC (2012). Heterokromatin dan penentuan posisi gen: di dalam, di luar, di sisi mana saja?. Kromosom , 121 (6), 555–563.
- Lewin, B. (2008). Gen IX . Penerbit Jones dan Bartlett.
- Tollefsbol, UNTUK (2011). Buku Pegangan Epigenetik . Pers Akademik.
- Wang, J., Jia, ST, & Jia, S. (2016). Wawasan Baru tentang Regulasi Heterochromatin. Tren genetika: TIG , 32 (5), 284-294.
- Zhang, P., Torres, K., Liu, X., Liu, CG, & Pollock, RE (2016). Ikhtisar Protein Pengatur Kromatin dalam Sel. Ilmu protein & peptida saat ini , 17 (5), 401-410.
