
enzim restriksi yang endonuklease yang digunakan oleh archaea dan tertentu bakteri untuk menghambat atau “membatasi” penyebaran virus dalam. Mereka sangat umum pada bakteri dan merupakan bagian dari sistem pertahanan mereka terhadap DNA asing yang dikenal sebagai sistem restriksi / modifikasi.
Enzim-enzim ini mengkatalisis pembelahan DNA pita ganda di lokasi tertentu, dapat direproduksi dan tanpa menggunakan energi tambahan. Sebagian besar membutuhkan keberadaan kofaktor seperti magnesium atau kation divalen lainnya, meskipun beberapa juga membutuhkan ATP atau S-adenosil metionin.
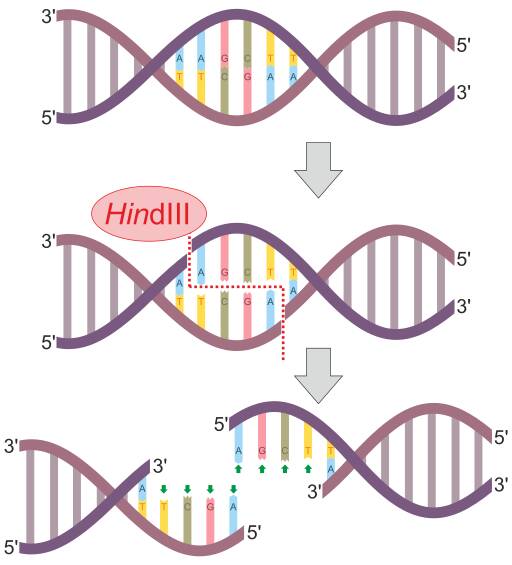
Skema reaksi enzim restriksi HindIII (Sumber: Helixitta [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)] melalui Wikimedia Commons)
Endonuklease restriksi ditemukan pada tahun 1978 oleh Daniel Nathans, Arber Werner, dan Hamilton Smith, yang menerima Hadiah Nobel dalam bidang kedokteran untuk penemuan mereka. Nama mereka umumnya berasal dari organisme tempat mereka pertama kali diamati.
Enzim tersebut banyak digunakan dalam pengembangan metode kloning DNA dan biologi molekuler lainnya serta strategi rekayasa genetika. Karakteristik pengenalan sekuens spesifik mereka dan kemampuan untuk memotong sekuens yang dekat dengan situs pengenalan menjadikannya alat yang ampuh dalam eksperimen genetik.
Fragmen yang dihasilkan oleh enzim restriksi yang bekerja pada molekul DNA tertentu dapat digunakan untuk membuat ulang “peta” molekul asli dengan menggunakan informasi tentang tempat enzim memotong DNA.
Beberapa enzim restriksi mungkin memiliki tempat pengenalan yang sama pada DNA, tetapi mereka tidak harus memotongnya dengan cara yang sama. Jadi, ada enzim yang memotong ujung tumpul dan enzim yang memotong ujung kohesif, yang memiliki aplikasi berbeda dalam biologi molekuler.
Saat ini ada ratusan enzim restriksi yang tersedia secara komersial, yang ditawarkan oleh rumah komersial yang berbeda; Enzim-enzim ini berfungsi sebagai gunting molekuler “khusus” untuk tujuan yang berbeda.
Indeks artikel
Fitur
Enzim restriksi memenuhi fungsi kebalikan dari polimerase, karena mereka menghidrolisis atau memutuskan ikatan ester dalam ikatan fosfodiester antara nukleotida yang berdekatan dalam rantai nukleotida.
Dalam biologi molekuler dan rekayasa genetika mereka banyak digunakan alat untuk konstruksi ekspresi dan vektor kloning, serta untuk identifikasi urutan tertentu. Mereka juga berguna untuk konstruksi genom rekombinan dan memiliki potensi bioteknologi yang besar.
Kemajuan terbaru dalam terapi gen membuat penggunaan enzim restriksi saat ini untuk pengenalan gen tertentu ke dalam vektor yang merupakan kendaraan untuk pengangkutan gen tersebut ke dalam sel hidup, dan yang mungkin memiliki kemampuan untuk menyisipkan ke dalam genom seluler untuk melakukan perubahan permanen.
Mekanisme aksi
Enzim restriksi dapat mengkatalisis pembelahan DNA pita ganda, meskipun beberapa mampu mengenali urutan DNA pita tunggal dan bahkan RNA . Pemotongan terjadi setelah pengenalan urutan.
Mekanisme aksi terdiri dari hidrolisis ikatan fosfodiester antara gugus fosfat dan deoksiribosa di tulang punggung setiap untai DNA. Banyak enzim mampu memotong di tempat yang sama yang mereka kenali, sementara yang lain memotong antara 5 dan 9 pasangan basa sebelum atau sesudahnya.
Enzim-enzim ini biasanya memotong pada ujung 5 ‘dari gugus fosfat, sehingga menimbulkan fragmen DNA dengan ujung fosforil 5’ dan ujung hidroksil 3 ‘terminal.
Karena protein tidak bersentuhan langsung dengan situs pengenalan dalam DNA, mereka harus ditranslokasikan secara berurutan sampai situs spesifik tercapai, mungkin melalui mekanisme “geser” pada untai DNA.
Selama pembelahan enzimatik, ikatan fosfodiester dari masing-masing untai DNA diposisikan dalam salah satu situs aktif enzim restriksi. Ketika enzim meninggalkan situs pengenalan dan pembelahan, ia melakukannya melalui asosiasi transien non-spesifik.
Jenis
Lima jenis enzim restriksi saat ini dikenal. Berikut adalah deskripsi singkat dari masing-masing:
Enzim restriksi tipe I
Enzim-enzim ini adalah protein pentamerik besar dengan tiga subunit, satu untuk restriksi, satu untuk metilasi, dan satu untuk pengenalan urutan dalam DNA. Endonuklease ini merupakan protein multifungsi yang mampu mengkatalisis reaksi restriksi dan modifikasi, memiliki aktivitas ATPase dan juga DNA topoisomerase.
Enzim jenis ini adalah endonuklease pertama yang ditemukan, mereka pertama kali dimurnikan pada 1960-an dan telah dipelajari secara mendalam sejak saat itu.
Enzim tipe I tidak banyak digunakan sebagai alat bioteknologi, karena situs pembelahan dapat berada pada jarak variabel hingga 1.000 pasangan basa dari situs pengenalan, yang membuatnya tidak dapat diandalkan dalam hal reproduktifitas eksperimental.
Enzim restriksi tipe II
Mereka adalah enzim yang terdiri dari homodimer atau tetramer yang memotong DNA di lokasi yang ditentukan antara 4 dan 8 bp panjangnya. Situs pembelahan ini biasanya palindromik, yaitu, mereka mengenali urutan yang dibaca dengan cara yang sama di kedua arah.
Banyak enzim restriksi tipe II pada bakteri memotong DNA ketika mereka mengenali karakter asingnya, karena ia tidak memiliki modifikasi khas yang seharusnya dimiliki DNA-nya sendiri.
Ini adalah enzim restriksi yang paling sederhana karena mereka tidak memerlukan kofaktor selain magnesium (Mg +) untuk mengenali dan memotong urutan DNA.
Ketepatan enzim restriksi tipe II dalam mengenali dan memotong sekuens sederhana dalam DNA pada posisi yang tepat menjadikannya salah satu yang paling banyak digunakan dan sangat diperlukan di sebagian besar cabang biologi molekuler.
Dalam kelompok enzim restriksi tipe II ada beberapa subkelas yang diklasifikasikan menurut sifat-sifat tertentu yang unik untuk masing-masing subkelas. Klasifikasi enzim ini dilakukan dengan menambahkan huruf abjad, dari A sampai Z mengikuti nama enzim.
Beberapa subclass yang terkenal karena kegunaannya adalah:
Subkelas IIA
Mereka adalah dimer dari subunit yang berbeda. Mereka mengenali urutan asimetris dan digunakan sebagai prekursor ideal untuk generasi enzim pemotongan.
Subkelas IIB
Mereka terdiri dari satu atau lebih dimer dan memotong DNA di kedua sisi urutan pengenalan. Mereka memotong kedua untai DNA dengan interval pasangan basa di depan situs pengenalan.
Subkelas IIC
Enzim jenis ini merupakan polipeptida dengan fungsi pembelahan dan modifikasi untai DNA. Enzim ini memotong kedua untai secara asimetris.
Subkelas IIE
Enzim dari subkelas ini adalah yang paling banyak digunakan dalam rekayasa genetika. Mereka memiliki situs katalitik dan umumnya membutuhkan efektor alosterik. Enzim-enzim ini perlu berinteraksi dengan dua salinan urutan pengenalannya untuk melakukan pembelahan yang efisien. Dalam subkelas ini adalah enzim EcoRII dan EcoRI.
Enzim restriksi tipe III
Endonuklease restriksi tipe III hanya terdiri dari dua subunit, satu bertanggung jawab untuk pengenalan dan modifikasi DNA, sementara yang lain bertanggung jawab untuk pembelahan urutan.
Enzim-enzim ini membutuhkan dua kofaktor untuk fungsinya: ATP dan magnesium. Enzim restriksi jenis ini memiliki dua situs pengenalan asimetris, mentranslokasi DNA dengan cara yang bergantung pada ATP dan memotongnya antara 20 hingga 30 bp yang berdekatan dengan situs pengenalan.
Enzim restriksi tipe IV
Enzim tipe IV mudah diidentifikasi karena mereka memotong DNA dengan tanda metilasi, mereka terdiri dari beberapa subunit berbeda yang bertanggung jawab untuk mengenali dan memotong urutan DNA. Enzim ini menggunakan GTP dan magnesium divalen sebagai kofaktor.
Situs pembelahan spesifik termasuk untai nukleotida dengan residu sitosin termetilasi atau hidroksimetilasi pada satu atau kedua untai asam nukleat.
Enzim restriksi tipe V
Klasifikasi ini mengelompokkan enzim tipe CRISPER-Cas, yang mengidentifikasi dan memotong urutan DNA spesifik dari organisme penyerang. Enzim Cas menggunakan untai RNA pemandu yang disintesis dari CRISPER untuk mengenali dan menyerang organisme yang menyerang.
Enzim yang diklasifikasikan sebagai tipe V adalah polipeptida yang disusun oleh enzim tipe I, II dan II. Mereka dapat memotong bagian DNA dari hampir semua organisme dan dengan rentang panjang yang lebar. Fleksibilitas dan kemudahan penggunaannya menjadikan enzim ini salah satu alat yang paling banyak digunakan dalam rekayasa genetika saat ini, bersama dengan enzim tipe II.
Contoh
Enzim restriksi telah digunakan untuk mendeteksi polimorfisme DNA, terutama dalam studi genetik populasi dan studi evolusi menggunakan DNA mitokondria, untuk memperoleh informasi tentang tingkat substitusi nukleotida.
Saat ini, vektor yang digunakan untuk transformasi bakteri untuk berbagai tujuan memiliki situs multiklon di mana situs pengenalan untuk beberapa enzim restriksi ditemukan.
Di antara enzim-enzim ini yang paling populer adalah EcoRI, II, III, IV dan V, diperoleh dan dideskripsikan untuk pertama kalinya dari E. coli ; HindIII dari H. influenzae dan BamHI dari B. amyloliquefaciens.
Referensi
- Bickle, TA, & Kruger, DH (1993). Biologi Pembatasan DNA. Ulasan Mikrobiologi , 57 (2), 434–450.
- Boyaval, P., Moineau, S., Romero, DA, & Horvath, P. (2007). CRISPR Memberikan resistensi yang didapat terhadap virus pada prokariota. Sains , 315 (Maret), 1709–1713.
- Goodsell, D. (2002). Perspektif molekuler: Pembatasan Endonuklease. Dasar-dasar Sel Induk Pengobatan Kanker , 20 , 190-191.
- Halford, SE (2001). Melompat, melompat dan mengulang dengan enzim restriksi. Transaksi Masyarakat Biokimia , 29 , 363-373.
- Jeltsch, A. (2003). Pemeliharaan identitas spesies dan pengendalian spesiasi bakteri: fungsi baru untuk sistem restriksi / modifikasi? Gen , 317 , 13-16.
- Krebs, J., Goldstein, E., & Kilpatrick, S. (2018). Gen XII Lewin (12 ed.). Burlington, Massachusetts: Pembelajaran Jones & Bartlett.
- Li, Y., Pan, S., Zhang, Y., Ren, M., Feng, M., Peng, N.,… Dia, Q. (2015). Memanfaatkan sistem CRISPR-Cas Tipe I dan Tipe III untuk pengeditan genom. Penelitian Asam Nukleat , 1–12.
- Loenen, WAM, Dryden, DTF, Raleigh, EA, & Wilson, GG (2013). Enzim restriksi tipe I dan kerabatnya. Penelitian Asam Nukleat , 1–25.
- Nathans, D., & Smith, HO (1975). Pembatasan Endonuklease dalam analisis dan restrukturisasi molekul DNA. annu. Pdt. Biokimia. , 273–293.
- Nei, M., & Tajima, F. (1981). Polimorfisme DNA terdeteksi oleh endonuklease restriksi. Genetika , 145-163.
- Pingoud, A., Fuxreiter, M., Pingoud, V., & Wende, W. (2005). Cellular and Molecular Life Sciences Tipe II restriksi endonuklease: struktur dan mekanisme. CMLS Seluler dan Molekular Biologi , 62 , 685-707.
- Roberts, R. (2005). Bagaimana enzim restriksi menjadi pekerja keras biologi molekuler. PNAS , 102 (17), 5905-5908.
- Roberts, RJ, & Murray, K. (1976). endonuklease restriksi. Ulasan Kritis dalam Biokimia , (November), 123-164.
- Stoddard, BL (2005). Menyebutkan struktur dan fungsi endonuklease. Ulasan Triwulanan Biofisika , 1–47.
- Tock, MR, & Dryden, DTF (2005). Biologi pembatasan dan anti-pembatasan. Opini Saat Ini dalam Mikrobiologi , 8 , 466-472. https://doi.org/10.1016/j.mib.2005.06.003
- Wilson, GG, & Murray, NE (1991). Sistem Pembatasan dan Modifikasi. annu. Pdt. , 25 , 585-627.
- Wu, Z., & Mou, K. (2016). Wawasan genom ke virulensi Campylobacter jejuni dan genetika populasi. menginfeksi Dis. terjemahan Med. , 2 (3), 109–119.
- Yuan, R. (1981). Struktur dan Mekanisme Endonuklease Pembatasan Multifungsi. annu. Pdt. Biokimia. , 50 , 285-315.
