
sphingomyelin adalah yang paling sphingolipids melimpah di jaringan hewan: diketahui terjadi di semua membran sel dipelajari untuk saat ini. Ini memiliki kesamaan struktural dengan fosfatidilkolin dalam hal kelompok kepala kutub, itulah sebabnya ia juga diklasifikasikan sebagai fosfolipid (fosfosfingolipid).
Pada tahun 1880-an, ilmuwan Johann Thudichum mengisolasi komponen lipid yang larut dalam eter dari jaringan otak dan menamakannya sphingomyelin. Kemudian, pada tahun 1927, struktur sphingolipid ini dilaporkan sebagai N -acyl-sphingosine-1-phosphocholine.
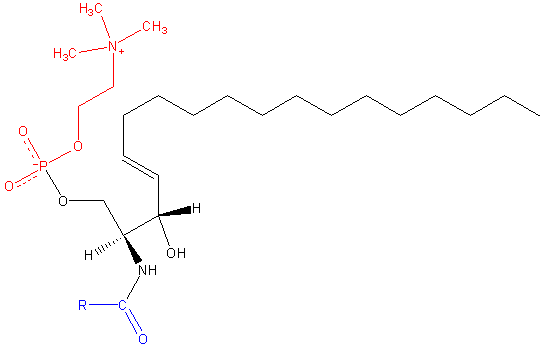
Struktur sphingomyelin (Sumber: Jag123 di Wikipedia bahasa Inggris. [Domain publik], melalui Wikimedia Commons)
Seperti sphingolipid lainnya, sphingomyelin memiliki fungsi struktural dan sinyal sel, dan terutama berlimpah di jaringan saraf, khususnya di mielin, selubung yang menutupi dan mengisolasi akson neuron tertentu.
Distribusinya telah dipelajari melalui fraksinasi subseluler dan eksperimen degradasi enzimatik dengan sphingomyelinase, dan hasilnya menunjukkan bahwa lebih dari setengah sphingomyelin dalam sel eukariotik ditemukan di membran plasma. Namun, ini tergantung pada jenis sel. Dalam fibroblas , misalnya, itu menyumbang hampir 90% dari total lipid.
Disregulasi proses sintesis dan metabolisme lipid ini mengarah pada perkembangan patologi kompleks atau lipidosis. Contohnya adalah penyakit Niemann-Pick herediter, yang ditandai dengan hepatosplenomegali dan disfungsi neurologis progresif.
Indeks artikel
Struktur
Sphingomyelin adalah molekul amfipatik yang terdiri dari kepala kutub dan dua ekor apolar. Gugus kepala kutub adalah molekul fosfokolin, sehingga mungkin tampak mirip dengan gliserofosfolipid fosfatidilkolin (PC). Namun, ada perbedaan substansial mengenai wilayah antarmuka dan hidrofobik antara kedua molekul ini.
Basa yang paling umum dalam molekul sphingomyelin mamalia adalah ceramide, terdiri dari sphingosine (1,3-dihydroxy-2-amino-4-octadecene), yang memiliki ikatan rangkap trans antara karbon pada posisi 4 dan 5 dari rantai hidrokarbon. Turunan jenuhnya, sphinganine, juga umum, tetapi ditemukan pada tingkat yang lebih rendah.
Panjang ekor hidrofobik sphingomyelin berkisar antara 16 sampai 24 atom karbon, dan komposisi asam lemak bervariasi tergantung pada jaringan.
sphingomyelins dari materi putih dari otak manusia , misalnya, memiliki asam nervonic, orang-orang dari materi abu-abu mengandung asam stearat terutama, dan bentuk lazim di trombosit adalah arakidonat.
Umumnya ada perbedaan panjang antara dua rantai asam lemak sphingomyelin, yang tampaknya mendukung fenomena “interdigitasi” antara hidrokarbon di lapisan tunggal yang berlawanan. Ini memberikan membran dengan stabilitas khusus dan sifat tertentu dibandingkan dengan membran lain yang lebih miskin dalam sphingolipid ini.
Di wilayah antarmuka molekul, sphingomyelin memiliki gugus amida dan hidroksil bebas pada karbon 3, yang dapat berfungsi sebagai donor dan akseptor ikatan hidrogen untuk ikatan intra dan antar molekul, penting dalam definisi domain samping dan interaksi dengan berbagai jenis. dari molekul.
Fitur
-Sinyal
Produk metabolisme sphingosine -ceramide, sphingosine, sphingosine 1-phosphate dan diacylglycerol- adalah efektor seluler yang penting dan memberikannya peran dalam berbagai fungsi seluler, seperti apoptosis, perkembangan dan penuaan, pensinyalan sel, antara lain.
-Struktur
Berkat struktur “silindris” tiga dimensi dari sphingomyelin, lipid ini dapat membentuk domain membran yang lebih kompak dan teratur, yang memiliki implikasi fungsional penting dari sudut pandang protein, karena dapat membentuk domain spesifik untuk beberapa protein membran integral.
Dalam lipid “rakit” dan caveolae
Rakit lipid, fase membran atau domain mikro yang dipesan dari sphingolipid seperti sphingomyelin, beberapa gliserofosfolipid dan kolesterol, mewakili platform yang stabil untuk asosiasi protein membran dengan berbagai fungsi (reseptor, transporter, dll.).
Caveolae adalah invaginasi membran plasma yang merekrut protein dengan jangkar GPI dan juga kaya akan sphingomyelin.
Sehubungan dengan kolesterol
Kolesterol, karena kekakuan strukturalnya, secara signifikan mempengaruhi struktur membran sel, terutama dalam aspek yang berkaitan dengan fluiditas, oleh karena itu dianggap sebagai unsur penting.
Karena sphingomyelin memiliki donor dan akseptor ikatan hidrogen, mereka diyakini mampu membentuk interaksi yang lebih “stabil” dengan molekul kolesterol. Untuk alasan ini, dikatakan bahwa ada korelasi positif antara kadar kolesterol dan sphingomyelin pada membran.
Perpaduan
Sintesis sfingomielin terjadi di kompleks Golgi , di mana seramida yang diangkut dari retikulum endoplasma (RE) dimodifikasi oleh transfer molekul fosfokolin dari fosfatidilkolin, dengan pelepasan molekul diasilgliserol secara bersamaan. Reaksi dikatalisis oleh SM sintase (ceramide: phosphatidylcholine phosphocholine transferase).
Ada juga jalur lain dari produksi sphingomyelin yang dapat terjadi dengan mentransfer phosphoethanolamine dari phosphatidylethanolamine (PE) ke ceramide, dengan metilasi phosphoethanolamine berikutnya. Ini dianggap sangat penting di beberapa jaringan saraf yang kaya PE.
Sfingomielin sintase ditemukan di sisi luminal membran kompleks Golgi, yang konsisten dengan lokasi ekstrasitoplasma sfingomielin di sebagian besar sel.
Karena karakteristik kelompok kutub sfingomielin dan tidak adanya translokase spesifik, orientasi topologi lipid ini bergantung pada enzim sintase.
Metabolisme
Degradasi sphingomyelin dapat terjadi pada membran plasma dan lisosom. Hidrolisis lisosom menjadi seramida dan fosfokolin bergantung pada sfingomielinase asam, suatu glikoprotein lisosom larut yang aktivitasnya memiliki pH optimal sekitar 4,5.
Hidrolisis dalam membran plasma dikatalisis oleh sphingomyelinase yang bekerja pada pH 7,4 dan membutuhkan ion magnesium atau mangan divalen untuk operasinya. Enzim lain yang terlibat dalam metabolisme dan daur ulang sphingomyelin ditemukan di berbagai organel yang terhubung satu sama lain melalui jalur transportasi vesikular.
Referensi
- Barenholz, Y., & Thompson, TE (1999). Sphingomyelin: aspek biofisik. Kimia dan Fisika Lipid , 102 , 29-34.
- Kanfer, J., & Hakomori, S. (1983). Biokimia Sphingolipid . (D. Hanahan, Ed.), Buku Pegangan Penelitian Lipid 3 (Edisi ke-1). Pers Pleno.
- Koval, M., & Pagano, R. (1991). Transportasi intraseluler dan metabolisme sphingomyelin. Biokimik , 1082 , 113-125.
- Lodish, H., Berk, A., Kaiser, CA, Krieger, M., Bretscher, A., Ploegh, H., Martin, K. (2003). Biologi Sel Molekuler (edisi ke-5). Freeman, WH & Perusahaan.
- Millat, G., Chikh, K., Naureckiene, S., Sleat, DE, Fensom, AH, Higaki, K.,… Vanier, MT (2001). Penyakit Niemann-Pick Tipe C: Spektrum Mutasi HE1 dan Korelasi Genotipe / Fenotipe dalam Grup NPC2. Am.J.Hum. gen. , 69 , 1013-1021.
- Ramstedt, B., & Slotte, P. (2002). Sifat membran sfingomielin. Surat FEBS , 531 , 33–37.
- Slotte, P. (1999). Sphingomyelin – interaksi kolesterol dalam membran biologis dan caral. Kimia dan Fisika Lipid , 102 , 13-27.
- Vance, JE, & Vance, DE (2008). Biokimia lipid, lipoprotein dan membran. Dalam Biokimia Komprehensif Baru Vol.36 (edisi ke-4). lain.
