
mikrofilamen atau aktin filamen, adalah salah satu dari tiga komponen utama dari sitoskeleton sel eukariotik (mikrofilamen, mikrotubulus dan filamen menengah) dan terdiri dari filamen kecil dari protein yang disebut aktin (aktin polimer).
Pada eukariota, gen yang mengkode mikrofilamen aktin sangat terkonservasi di semua organisme, itulah sebabnya mereka sering digunakan sebagai penanda molekuler untuk berbagai jenis studi.
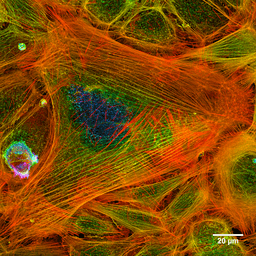
Foto filamen aktin sel yang diwarnai (Sumber: Howard Vindin [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)] melalui Wikimedia Commons)
Mikrofilamen didistribusikan ke seluruh sitosol, tetapi terutama berlimpah di wilayah di bawah membran plasma, di mana mereka membentuk jaringan kompleks dan berasosiasi dengan protein khusus lainnya untuk membentuk sitoskeleton.
Jaringan mikrofilamen dalam sitoplasma sel mamalia dikodekan oleh dua dari enam gen yang dijelaskan untuk aktin, yang terlibat dalam dinamika mikrofilamen dan bahkan sangat penting selama diferensiasi sel induk.
Banyak penulis setuju bahwa mikrofilamen adalah protein yang paling beragam, serbaguna, dan penting dalam sitoskeleton sebagian besar sel eukariotik, dan penting untuk diingat bahwa ini tidak ditemukan pada mikroorganisme prokariotik.
Dalam jenis sel ini, di sisi lain, ada filamen yang homolog dengan mikrofilamen, tetapi terdiri dari protein lain: protein MreB.
Gen yang mengkode protein ini saat ini dianggap sebagai gen leluhur yang mungkin untuk aktin eukariotik. Namun, urutan homologi asam amino yang membentuk protein MreB hanya 15% sehubungan dengan urutan aktin.
Karena mereka adalah bagian mendasar dari sitoskeleton, setiap cacat fenotipik di kedua mikrotubulus dan filamen menengah dan mikrofilamen aktin (sitoskeleton) dapat menyebabkan patologi seluler dan sistemik yang berbeda.
Indeks artikel
Karakteristik dan struktur
Mikrofilamen terdiri dari monomer protein dari keluarga aktin, yang merupakan protein kontraktil yang sangat melimpah dalam sel eukariotik, karena mereka juga berpartisipasi dalam kontraksi otot.
Filamen ini memiliki diameter antara 5 dan 7 nm, sehingga mereka juga dikenal sebagai filamen tipis dan terdiri dari dua bentuk aktin: bentuk globular (G aktin) dan bentuk filamen (F aktin).
Protein yang berpartisipasi dalam sitoskeleton dikenal sebagai dan aktin, sedangkan yang berpartisipasi dalam kontraksi biasanya -aktin.
Proporsi aktin globular dan aktin berfilamen dalam sitosol tergantung pada kebutuhan seluler, karena mikrofilamen adalah struktur yang sangat bervariasi dan serbaguna, yang terus tumbuh dan memendek melalui polimerisasi dan depolimerisasi.
G aktin adalah protein globular kecil, terdiri dari hampir 400 asam amino dan dengan berat molekul sekitar 43 kDa .
Monomer G-aktin yang membentuk mikrofilamen disusun dalam bentuk untai heliks, karena masing-masing mengalami putaran ketika dikaitkan dengan yang berikutnya.
G aktin berasosiasi dengan satu molekul Ca2 + dan satu lagi dari ATP, yang menstabilkan bentuk globularnya; sedangkan F-aktin diperoleh setelah hidrolisis fosfat terminal molekul ATP menjadi G-aktin, yang berkontribusi pada polimerisasi.
Organisasi
Filamen aktin dapat diatur menjadi “bundel” atau “jaringan” yang memiliki fungsi berbeda di dalam sel. Bundel membentuk struktur paralel yang dihubungkan oleh jembatan silang yang cukup kaku.
Jaringan, di sisi lain, adalah struktur yang lebih longgar, seperti jerat tiga dimensi dengan sifat gel semi-padat.
Ada banyak protein yang berasosiasi dengan filamen aktin atau mikrofilamen dan dikenal sebagai ABP ( protein pengikat aktin ), yang memiliki situs spesifik untuk ini.
Banyak dari protein ini memungkinkan mikrofilamen untuk berinteraksi dengan dua komponen lain dari sitoskeleton: mikrotubulus dan filamen menengah, serta dengan komponen lain di permukaan bagian dalam membran plasma.
Protein lain yang berinteraksi dengan mikrofilamen termasuk lamina nuklir dan spektrin (dalam sel darah merah).
Bagaimana filamen aktin terbentuk?
Karena monomer aktin globular selalu mengikat dengan cara yang sama, berorientasi pada arah yang sama, mikrofilamen memiliki polaritas yang pasti, dengan dua ujung: satu “lebih” dan satu “kurang”.
Polaritas filamen ini sangat penting, karena mereka tumbuh jauh lebih cepat pada ujung positifnya, di mana monomer G-aktin baru ditambahkan.

Representasi grafis dari pembentukan mikrofilamen aktin (Sumber: karya turunan: Retama (bicara) Thin_filament_formation.svg: Mikael Häggström [Domain publik] melalui Wikimedia Commons)
Hal pertama yang terjadi selama polimerisasi filamen aktin adalah proses yang dikenal sebagai “nukleasi”, yang terdiri dari asosiasi tiga monomer protein.
Monomer baru ditambahkan ke trimer ini di kedua ujungnya, sehingga filamen tumbuh. Monomer G-aktin mampu menghidrolisis ATP dengan setiap pengikatan, yang berimplikasi pada laju polimerisasi, karena gugus aktin-ATP terdisosiasi dengan kesulitan yang lebih besar daripada gugus aktin-ADP.
ATP tidak diperlukan untuk polimerisasi dan peran spesifik hidrolisisnya belum dijelaskan.
Beberapa penulis menganggap bahwa, karena peristiwa polimerisasi aktin dapat dibalik dengan cepat, ATP yang terkait dengan proses ini dapat mewakili hingga 40% dari total pergantian seluler dari molekul energik ini.
Peraturan
Baik polimerisasi filamen aktin dan depolimerisasinya adalah proses yang sangat diatur oleh serangkaian protein spesifik, yang bertanggung jawab untuk recaraling filamen.
Contoh protein yang mengatur depolimerisasi adalah faktor depolimerisasi aktin cofilin. Protein lain, profilin, memiliki fungsi yang berlawanan, karena merangsang asosiasi monomer (dengan merangsang pertukaran ADP untuk ATP).
Fitur
Mikrofilamen berinteraksi dengan filamen miosin yang berasosiasi dengan protein transmembran yang memiliki domain di sitosol dan domain lain di luar sel, sehingga berpartisipasi dalam proses mobilitas sel.
Mikrofilamen yang terkait dengan membran plasma ini memediasi berbagai respons seluler terhadap berbagai kelas rangsangan. Misalnya, adhesi sel dalam jaringan epitel didorong oleh protein transmembran yang dikenal sebagai cadherin, yang berinteraksi dengan mikrofilamen untuk merekrut faktor respons.
Filamen aktin berinteraksi dengan filamen perantara untuk menyebabkan rangsangan ekstraseluler ditransmisikan ke lokasi utama seperti ribosom dan kromosom di dalam nukleus.
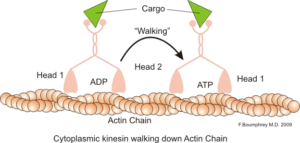
Representasi fungsi motorik intraseluler mikrofilamen aktin (Sumber: Boumphreyfr [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)] melalui Wikimedia Commons)
Fungsi klasik dan banyak dipelajari dari mikrofilamen adalah kemampuannya untuk membentuk “jembatan”, “rel” atau “jalan raya” untuk pergerakan protein motorik miosin I, yang mampu memuat vesikel transpor dari organel ke plasma membran di jalur sekretori.
Mikrofilamen juga berinteraksi dengan miosin II untuk membentuk cincin kontraktil yang terbentuk selama sitokinesis, tepatnya pada tahap terakhir pembelahan sel di mana sitosol dipisahkan dari sel induk dan sel anak.
Secara umum, mikrofilamen F-aktin memodulasi distribusi beberapa organel seperti kompleks Golgi, retikulum endoplasma, dan mitokondria. Selain itu, mereka juga berpartisipasi dalam penentuan posisi spasial mRNA sehingga dibaca oleh ribosom.
Seluruh rangkaian mikrofilamen seluler, terutama yang terkait erat dengan membran plasma, berpartisipasi dalam pembentukan membran sel bergelombang yang memiliki gerakan aktif konstan.
Mereka juga terlibat dalam pembentukan mikrovili dan benjolan umum lainnya pada permukaan banyak sel.
Contoh fungsi hati
Mikrofilamen berpartisipasi dalam proses sekresi empedu di hepatosit (sel hati) dan juga dalam gerakan peristaltik (kontraksi terkoordinasi) kanalikuli hati.
Mereka berkontribusi pada diferensiasi domain membran plasma berkat hubungannya dengan unsur sitosol yang berbeda dan kontrol yang mereka berikan atas topografi unsur intraseluler ini.
Patologi terkait
Ada beberapa penyakit yang berhubungan dengan cacat primer pada struktur atau dengan protein pengatur dan enzim dalam sintesis mikrofilamen, meskipun faktanya ini terlibat langsung dalam sejumlah besar fungsi.
Rendahnya tingkat penyakit dan malformasi dalam struktur utama mikrofilamen disebabkan oleh fakta bahwa, secara umum, ada banyak gen yang mengkode aktin dan protein pengaturnya, sebuah fenomena yang dikenal sebagai “redundansi genetik”.
Salah satu patologi yang paling banyak dipelajari adalah vitrifikasi oosit pada sitoskeleton mereka, di mana gangguan dalam jaringan mikrofilamen kortikal diamati, serta depolimerisasi dan disorganisasi mikrotubulus dari gelendong mitosis.
Secara umum, vitrifikasi ini menyebabkan dispersi kromosom, karena menyebabkan kekacauan dalam pemadatan semua kromatin.
Sel-sel yang memiliki organisasi dan proporsi mikrofilamen yang lebih besar dalam sitoskeletonnya adalah sel-sel otot lurik, oleh karena itu, sebagian besar patologi dikaitkan dengan kerusakan alat kontraktil.
Mikrofilamen yang rusak atau atipikal juga telah dikaitkan dengan penyakit tulang yang dikenal sebagai penyakit Paget.
Referensi
- Aguilar-Cuenca, R., Llorente-González, C., Vicente, C., & Vicente-Manzanares, M. (2017). Dinamika adhesi terkoordinasi mikrofilamen mendorong migrasi sel tunggal dan membentuk seluruh jaringan. F1000Penelitian , 6 .
- Dos Remedios, CG, Chhabra, D., Kekic, M., Dedova, IV, Tsubakihara, M., Berry, DA, & Nosworthy, NJ (2003). Protein pengikat aktin: regulasi mikrofilamen sitoskeletal. Tinjauan Fisiologis , 83 (2), 433-473.
- Guo, H., Fauci, L., Shelley, M., & Kanso, E. (2018). Bistabilitas dalam sinkronisasi mikrofilamen yang digerakkan. Jurnal Mekanika Fluida, 836, 304-323.
- Lanza, R., Langer, R., & Vacanti, JP (Eds.). (2011). Prinsip rekayasa jaringan. pers akademik.
- Robbins, J. (2017). Penyakit Sitoskeleton: Desminopathies. Pada Miopati Kardioskeletal pada Anak dan Dewasa Muda (hlm. 173-192). Pers Akademik.
