
sphingolipids adalah salah satu dari tiga keluarga besar lipid dalam membran biologis. Seperti gliserofosfolipid dan sterol, mereka adalah molekul amfipatik dengan daerah kutub hidrofilik dan daerah apolar hidrofobik.
Mereka pertama kali dijelaskan pada tahun 1884 oleh Johann LW Thudichum, yang menggambarkan tiga sphingolipids (sphingomyelin, cerebrosides, dan cerebrosulfatide) yang termasuk dalam tiga kelas yang diketahui berbeda: phosphoesphingolipids, glikosfingolipid netral dan asam.
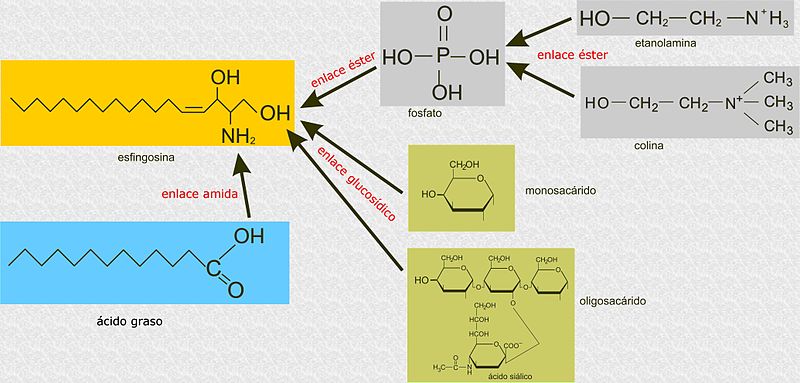
Alejandro Porto [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], melalui Wikimedia Commons
Tidak seperti gliserofosfolipid, sphingolipid tidak dibangun di atas molekul gliserol 3-fosfat sebagai kerangka utama, tetapi merupakan senyawa yang berasal dari sphingosine, suatu amino alkohol dengan rantai hidrokarbon panjang yang dihubungkan oleh ikatan amida.
Dalam hal kompleksitas dan keragaman, setidaknya 5 jenis basa yang berbeda diketahui untuk sphingolipid pada mamalia . Basa-basa ini dapat bergabung dengan lebih dari 20 jenis asam lemak yang berbeda, dengan panjang dan derajat kejenuhan yang bervariasi, selain banyaknya variasi gugus polar yang dapat terjadi.
Membran biologis memiliki sekitar 20% sphingolipid. Ini memiliki fungsi yang bervariasi dan penting dalam sel, dari struktural hingga transduksi sinyal, dan kontrol proses komunikasi seluler yang berbeda.
Distribusi molekul-molekul ini bervariasi tergantung pada fungsi organel di mana mereka ditemukan, tetapi biasanya konsentrasi sphingolipids jauh lebih tinggi di monolayer luar membran plasma daripada di monolayer dalam dan kompartemen lainnya.
Pada manusia setidaknya ada 60 spesies sphingolipid. Banyak dari mereka merupakan komponen penting dari membran sel saraf, sementara yang lain memainkan peran struktural penting atau berpartisipasi dalam transduksi sinyal, pengenalan, diferensiasi sel , patogenesis, kematian sel terprogram, antara lain.
Indeks artikel
Struktur
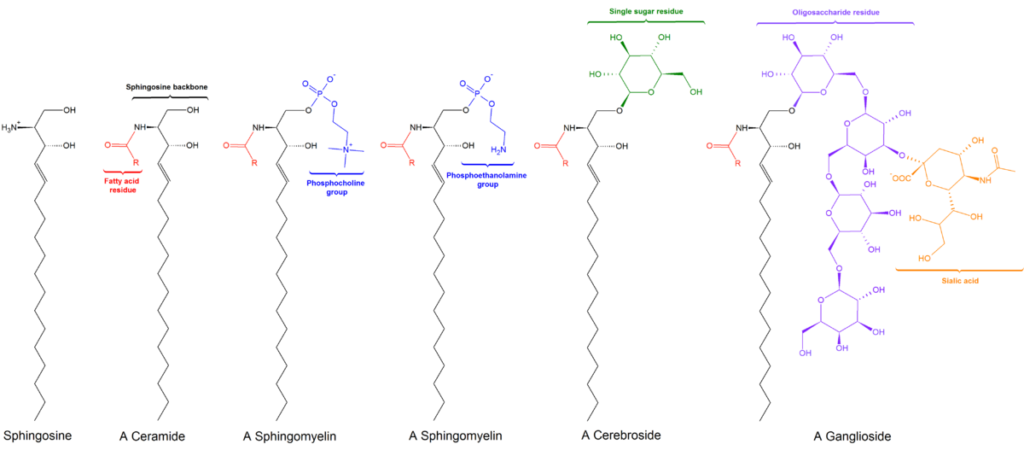
Struktur umum sphingolipids. LHcheM [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], dari Wikimedia Commons
Semua sphingolipid berasal dari L-serin, yang dikondensasi dengan asam lemak rantai panjang untuk membentuk basa sphingoid, juga dikenal sebagai basa rantai panjang (LCB).
Basa yang paling umum adalah sphinganine dan sphingosine, yang berbeda satu sama lain hanya dengan adanya ikatan rangkap trans antara karbon 4 dan 5 dari asam lemak sphingosine.
Karbon 1, 2, dan 3 dari sphingosine secara struktural analog dengan karbon gliserol dari gliserofosfolipid. Ketika asam lemak terikat pada karbon 2 sphingosine melalui ikatan amida, sebuah ceramide diproduksi, yang merupakan molekul yang sangat mirip dengan diasilgliserol dan mewakili sphingolipid yang paling sederhana.
Asam lemak rantai panjang yang membentuk daerah hidrofobik lipid ini bisa sangat beragam. Panjangnya bervariasi dari 14 hingga 22 atom karbon yang dapat memiliki derajat kejenuhan yang berbeda, biasanya antara karbon 4 dan 5.
Pada posisi 4 atau 6 mereka dapat memiliki gugus hidroksil dan ikatan rangkap pada posisi lain atau bahkan bercabang seperti gugus metil.
Karakteristik
Rantai asam lemak yang dihubungkan oleh ikatan amida ke seramida umumnya jenuh, dan cenderung lebih panjang daripada yang ditemukan dalam gliserofosfolipid, yang tampaknya penting untuk aktivitas biologisnya.
Ciri khas kerangka sphingolipid adalah bahwa mereka dapat memiliki muatan positif bersih pada pH netral, jarang terjadi di antara molekul lipid.
Namun, pKa dari gugus amino rendah terhadap amina sederhana, antara 7 dan 8, sehingga sebagian molekul tidak bermuatan pada pH fisiologis, yang dapat menjelaskan pergerakan “bebas” ini antara bilayer. .
Klasifikasi tradisional sphingolipids muncul dari beberapa modifikasi yang molekul ceramide dapat mengalami, terutama dalam hal substitusi kelompok kepala kutub.
Fitur
Sfingolipid sangat penting pada hewan , tumbuhan, dan jamur , serta pada beberapa organisme dan virus prokariotik .
-Fungsi struktural
Sphingolipids memodulasi sifat fisik membran, termasuk fluiditas, ketebalan, dan kelengkungannya. Memodulasi sifat-sifat ini juga memberi mereka pengaruh langsung pada organisasi spasial protein membran.
Dalam lipid “rakit”
Dalam membran biologis, domain mikro dinamis dengan fluiditas yang lebih sedikit dapat dideteksi, terdiri dari molekul kolesterol dan sphingolipid yang disebut rakit lipid.
Struktur ini terjadi secara alami dan terkait erat dengan protein integral, reseptor permukaan sel dan protein pensinyalan, pengangkut, dan protein lain dengan jangkar glikosilfosfatidilinositol (GPI).
-Fungsi sinyal
Mereka memiliki fungsi sebagai molekul pemberi sinyal yang bertindak sebagai pembawa pesan kedua atau sebagai ligan yang disekresikan untuk reseptor permukaan sel.
Sebagai utusan sekunder mereka dapat berpartisipasi dalam regulasi homeostasis kalsium, pertumbuhan sel, tumorigenesis dan penekanan apoptosis. Selanjutnya, aktivitas banyak protein membran integral dan perifer bergantung pada hubungannya dengan sphingolipids.
Banyak interaksi antara sel dan sel dengan lingkungannya bergantung pada paparan kelompok kutub yang berbeda dari sphingolipids ke permukaan luar membran plasma.
Pengikatan glikosfingolipid dan lektin sangat penting untuk asosiasi mielin dengan akson, adhesi neutrofil ke endotelium, dll.
Produk sampingan dari metabolisme Anda
Sphingolipid pensinyalan yang paling penting adalah basa rantai panjang atau sphingosine dan ceramide, serta turunannya yang terfosforilasi, seperti sphingosine 1-phosphate.
Produk metabolisme dari banyak sphingolipids mengaktifkan atau menghambat beberapa target hilir (protein kinase, fosfoprotein fosfatase, dan lain-lain), yang mengontrol perilaku seluler yang kompleks seperti pertumbuhan, diferensiasi, dan apoptosis.
-Sebagai reseptor di membran
Beberapa patogen menggunakan glikosfingolipid sebagai reseptor untuk memediasi masuknya mereka ke dalam sel inang atau untuk memberikan faktor virulensi kepada mereka.
Sphingolipids telah terbukti berpartisipasi dalam beberapa peristiwa seluler seperti sekresi, endositosis, kemotaksis, neurotransmisi, angiogenesis, dan peradangan.
Mereka juga terlibat dalam perdagangan membran, sehingga mempengaruhi internalisasi reseptor, pemesanan, pergerakan dan fusi vesikel sekretori dalam menanggapi rangsangan yang berbeda.
Kelompok sphingolipid
Ada tiga subkelas sphingolipid, semuanya berasal dari ceramide dan berbeda satu sama lain oleh kelompok kutub, yaitu: sphingomyelins, glikolipid, dan gangliosida.
Sfingomielin
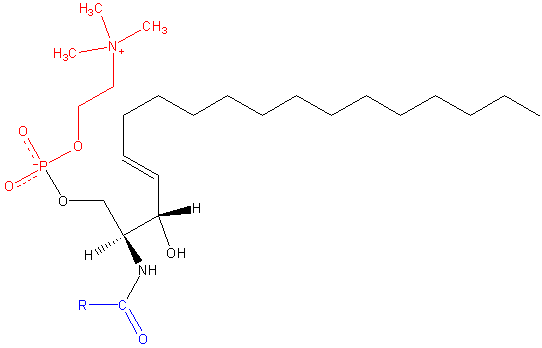
Sphingomilein. Hitam: sphingosine. Merah: fosfokolin. Biru: asam lemak.
Ini mengandung fosfokolin atau fosfoetanolamina sebagai gugus kepala polar, sehingga mereka diklasifikasikan sebagai fosfolipid bersama dengan gliserofosfolipid. Mereka menyerupai, tentu saja, fosfatidilkolin dalam struktur tiga dimensi dan sifat umum karena mereka tidak memiliki muatan di kepala kutubnya.
Mereka hadir dalam membran plasma sel hewan dan terutama berlimpah di mielin, selubung yang mengelilingi dan mengisolasi akson dari beberapa neuron.
Glikolipid netral atau glikosfingolipid (tanpa biaya)
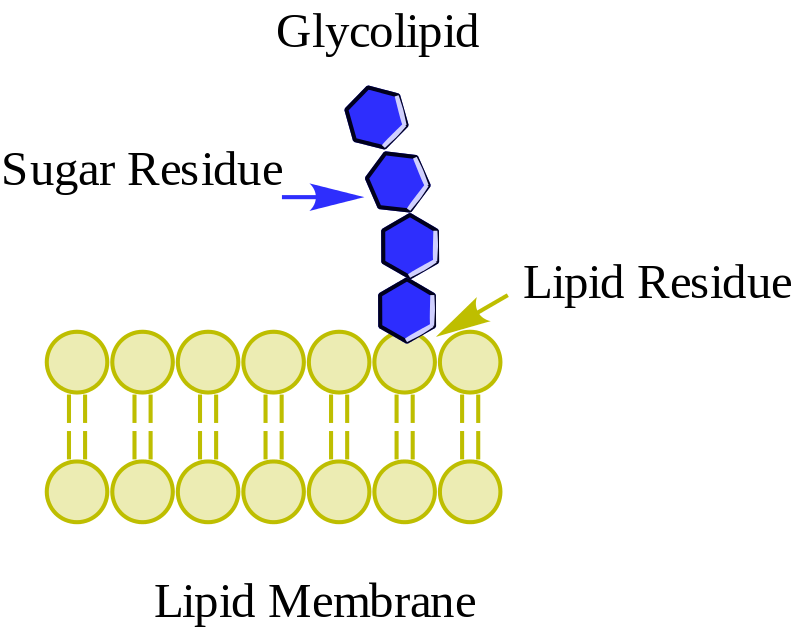
Glikolipid Wpcrosson [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)], dari Wikimedia Commons
Mereka ditemukan terutama pada permukaan luar membran plasma dan memiliki satu atau lebih gula sebagai gugus kepala polar yang terikat langsung pada hidroksil karbon 1 dari bagian seramida. Mereka tidak memiliki gugus fosfat. Karena pada pH 7 mereka tidak bermuatan, mereka disebut glikolipid netral.
Cerebrosides memiliki molekul gula tunggal yang melekat pada ceramide. Mereka yang mengandung galaktosa ditemukan dalam membran sel plasma jaringan non-saraf. Globosida adalah glikosfingolipid dengan dua atau lebih gula, biasanya D-glukosa, D-galaktosa, atau N-asetil-D-galaktosamin.
Gangliosida asam atau glycosphingolipids
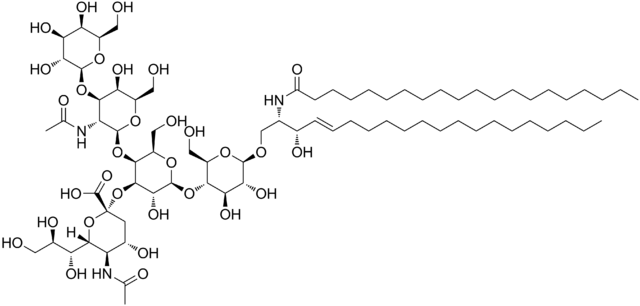
Struktur gangliosida GM1
Ini adalah sphingolipid yang paling kompleks. Mereka memiliki oligosakarida sebagai gugus kepala polar dan satu atau lebih residu asam N-asetilmuramat terminal, juga disebut asam sialat. Asam sialic memberi gangliosida muatan negatif pada pH 7, yang membedakannya dari glikosfingolipid netral.
Nomenklatur kelas sphingolipid ini tergantung pada jumlah residu asam sialat yang ada di bagian oligosakarida dari kepala kutub.
Perpaduan
Molekul basa rantai panjang, atau sphingosine, disintesis di retikulum endoplasma (ER) dan penambahan gugus polar ke kepala lipid ini terjadi kemudian di kompleks Golgi . Pada mamalia, beberapa sintesis sphingolipid juga dapat terjadi di mitokondria .
Setelah menyelesaikan sintesisnya di kompleks Golgi, sphingolipid diangkut ke kompartemen sel lain melalui mekanisme yang diperantarai vesikel.
Biosintesis sphingolipids terdiri dari tiga peristiwa mendasar: sintesis basa rantai panjang, biosintesis ceramide dengan penyatuan asam lemak melalui ikatan amida, dan akhirnya, pembentukan sphingolipid kompleks melalui penyatuan gugus polar. pada karbon 1 dari dasar sphingoid.
Selain sintesis de novo , sphingolipids juga dapat dibentuk oleh pergantian atau daur ulang basa rantai panjang dan ceramides, yang dapat memberi makan kumpulan sphingolipids.
Sintesis kerangka ceramide
Biosintesis ceramide, tulang punggung sphingolipids, dimulai dengan kondensasi dekarboksilat dari molekul palmitoyl-CoA dan L-serine. Reaksi dikatalisis oleh heterodimeric serin palmitoyl transferase (SPT), bergantung pada piridoksal fosfat dan produknya adalah 3-keto dihydrosphingosine.
Enzim ini dihambat oleh -halo-L-alanines dan L-cycloserines. Dalam ragi itu dikodekan oleh dua gen, sedangkan pada mamalia ada tiga gen untuk enzim ini. Situs aktif terletak di sisi sitoplasma retikulum endoplasma.
Peran enzim pertama ini dilestarikan di semua organisme yang dipelajari. Namun, ada beberapa perbedaan antara taksa yang berkaitan dengan lokasi subselular enzim: bakteri berada di sitoplasma, ragi, tumbuhan dan hewan berada di retikulum endoplasma.
3-ketosphinganine kemudian direduksi oleh 3-ketosphinganine reduktase yang bergantung pada NADPH untuk menghasilkan sphinganine. Dihydroceramide synthase (sphinganine N-acyl transferase) kemudian asetilasi sphinganine untuk menghasilkan dihydroceramide. Ceramide kemudian dibentuk oleh dihydroceramide desaturase/reductase, yang menyisipkan ikatan rangkap trans pada posisi 4-5.
Pada mamalia ada banyak isoform sintase ceramide, masing-masing mengikat rantai spesifik asam lemak ke basa rantai panjang. Oleh karena itu, ceramide sintase dan enzim lain, elongase, menyediakan sumber utama keragaman asam lemak dalam sphingolipids.
Pembentukan sphingolipid spesifik
Sfingomielin disintesis oleh transfer fosfokolin dari fosfatidilkolin ke ceramide, melepaskan diasilgliserol. Reaksi mengikat jalur sinyal sphingolipid dan gliserofosfolipid.
Phosphoethanolamine ceramide disintesis dari phosphatidylethanolamine dan ceramide dalam reaksi yang analog dengan sintesis sphingomyelin, dan setelah terbentuk dapat dimetilasi menjadi sphingomyelin. Inositol phosphate ceramides dibentuk oleh transesterifikasi dari phosphatidylinositol.
Glikosfingolipid dimodifikasi terutama di kompleks Golgi, di mana enzim glikosiltransferase spesifik berpartisipasi dalam penambahan rantai oligosakarida di wilayah hidrofilik tulang punggung ceramide.
Metabolisme
Degradasi sphingolipids dilakukan oleh enzim glukohidrolase dan sfingomielinase, yang bertugas menghilangkan modifikasi gugus polar. Di sisi lain, ceramidase meregenerasi basa rantai panjang dari ceramides.
Gangliosida terdegradasi oleh satu set enzim lisosom yang mengkatalisis penghapusan langkah demi langkah unit gula, akhirnya menghasilkan ceramide.
Jalur degradasi lain terdiri dari internalisasi sphingolipids dalam vesikel endositik yang dikirim kembali ke membran plasma atau diangkut ke lisosom di mana mereka didegradasi oleh hidrolase asam tertentu.
Tidak semua basa rantai panjang didaur ulang, retikulum endoplasma memiliki rute untuk degradasi terminalnya. Mekanisme degradasi ini terdiri dari fosforilasi daripada asilasi LCB, sehingga menimbulkan molekul pensinyalan yang dapat menjadi substrat larut untuk enzim liase yang membelah LCBs-fosfat untuk menghasilkan asilaldehida dan fosfoetanolamina.
Peraturan
Metabolisme lipid ini diatur pada berbagai tingkat, salah satunya adalah enzim yang bertanggung jawab atas sintesis, modifikasi pasca-translasi dan mekanisme alosteriknya.
Beberapa mekanisme pengaturan bersifat spesifik sel, baik untuk mengontrol momen perkembangan sel di mana mereka diproduksi atau sebagai respons terhadap sinyal tertentu.
Referensi
- Bartke, N., & Hannun, Y. (2009). Sphingolipids Bioaktif: Metabolisme dan Fungsi. Jurnal Penelitian Lipid , 50 , 19.
- Breslow, DK (2013). Homeostasis Sphingolipid di Retikulum Endoplasma dan Di Luarnya. Perspektif Pelabuhan Mata Air Dingin dalam Biologi , 5 (4), a013326.
- Futerman, AH, & Hannun, YA (2004). Kehidupan kompleks sphingolipid sederhana. Laporan EMBO , 5 (8), 777-782.
- Harrison, PJ, Dunn, T., & Campopiano, DJ (2018). Biosintesis sphingolipid pada manusia dan mikroba. Laporan Produk Alami , 35 (9), 921–954.
- Lahiri, S., & Futerman, AH (2007). Metabolisme dan fungsi sphingolipids dan glycosphingolipids. Ilmu Kehidupan Seluler dan Molekuler , 64 (17), 2270–2284.
- Lodish, H., Berk, A., Kaiser, CA, Krieger, M., Bretscher, A., Ploegh, H., Martin, K. (2003). Biologi Sel Molekuler (edisi ke-5). Freeman, WH & Perusahaan.
- Lucky, M. (2008). Biologi struktural membran: dengan dasar biokimia dan biofisika . Pers Universitas Cambridge. Diperoleh dari www.cambridge.org/9780521856553
- Merrill, AH (2011). Jalur metabolisme sphingolipid dan glycosphingolipid di era sphingolipidomik. Ulasan Kimia , 111 (10), 6387-6422.
- Nelson, DL, & Cox, MM (2009). Prinsip Biokimia Lehninger . Edisi Omega ( edisi ke-5).
- Vance, JE, & Vance, DE (2008). Biokimia lipid, lipoprotein dan membran. Dalam Biokimia Komprehensif Baru Vol.36 (edisi ke-4). lain.
