
nukleosom adalah unit kemasan dasar DNA pada organisme eukariotik. Oleh karena itu unsur kompresi terkecil untuk kromatin.
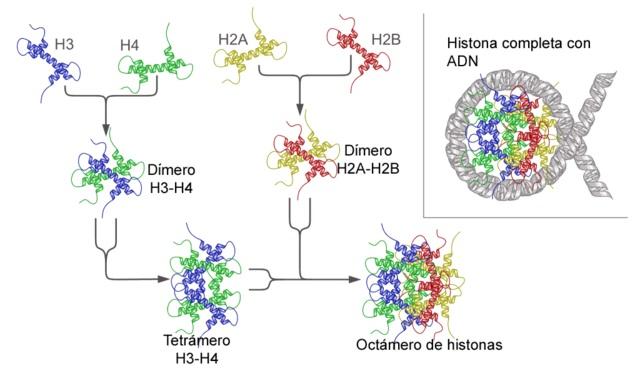
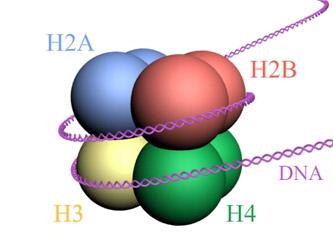
Nukleosom dibangun sebagai oktamer protein yang disebut histon, atau struktur berbentuk drum di mana sekitar 140 nt DNA dililit, membuat hampir dua putaran lengkap.
Struktur nukleosom
Selain itu, tambahan 40-80 nt DNA dianggap sebagai bagian dari nukleosom, dan itu adalah fraksi DNA yang memungkinkan kontinuitas fisik antara satu nukleosom dan yang lain dalam struktur kromatin yang lebih kompleks (seperti serat kromatin 30 nm).
Kode histone adalah salah satu unsur kontrol epigenetik pertama yang paling dipahami secara molekuler.
Indeks artikel
Fitur
Nukleosom memungkinkan:
- Pengemasan DNA agar sesuai dengan ruang inti yang terbatas.
- Mereka menentukan partisi antara kromatin yang diekspresikan (eukromatin) dan kromatin diam (heterokromatin).
- Mereka mengatur semua kromatin baik secara spasial maupun fungsional dalam nukleus.
- Mereka mewakili substrat modifikasi kovalen yang menentukan ekspresi, dan tingkat ekspresi, dari gen yang mengkode protein melalui apa yang disebut kode histone.
Komposisi dan struktur
Dalam pengertian yang paling dasar, nukleosom terdiri dari DNA dan protein. DNA dapat berupa DNA pita ganda apa pun yang ada dalam nukleus sel eukariotik , sedangkan protein nukleosomal semuanya termasuk dalam kumpulan protein yang disebut histon.
Histon adalah protein kecil dengan muatan residu asam amino basa yang tinggi; Hal ini memungkinkan untuk melawan muatan negatif DNA yang tinggi dan membangun interaksi fisik yang efisien antara dua molekul tanpa mencapai kekakuan ikatan kimia kovalen.
Histon membentuk oktamer mirip drum dengan dua salinan atau monomer dari masing-masing histon H2A, H2B, H3, dan H4. DNA membuat hampir dua putaran penuh pada sisi oktamer dan kemudian melanjutkan dengan sebagian kecil DNA penghubung yang berasosiasi dengan histon H1, untuk kembali membuat dua putaran penuh pada oktamer histon lainnya.
Himpunan oktamer, DNA terkait, dan DNA penghubungnya yang sesuai, adalah nukleosom.
Pemadatan kromatin
DNA genom terdiri dari molekul yang sangat panjang (lebih dari satu meter dalam kasus manusia, mengingat semua kromosom mereka), yang harus dipadatkan dan diatur dalam nukleus yang sangat kecil.
Langkah pertama dalam pemadatan ini dilakukan melalui pembentukan nukleosom. Dengan langkah ini saja, DNA dipadatkan sekitar 75 kali.
Ini menimbulkan serat linier dari mana tingkat pemadatan kromatin berikutnya dibangun: serat 30 nm, loop, dan loop loop.
Ketika sel membelah, baik dengan mitosis atau meiosis, tingkat pemadatan tertinggi adalah kromosom mitosis atau meiosis itu sendiri.
Kode histone dan ekspresi gen
Fakta bahwa oktamer histon dan DNA berinteraksi secara elektrostatis menjelaskan sebagian hubungan efektif mereka, tanpa kehilangan fluiditas yang diperlukan untuk membuat nukleosom menjadi unsur dinamis pemadatan dan dekompaksi kromatin.
Tetapi ada unsur interaksi yang lebih mengejutkan lagi: ujung N-terminal dari histone terekspos di luar interior oktamer yang lebih kompak dan inert.
Ujung-ujung ini tidak hanya berinteraksi secara fisik dengan DNA, tetapi juga mengalami serangkaian modifikasi kovalen di mana derajat pemadatan kromatin dan ekspresi DNA terkait akan bergantung.
Himpunan modifikasi kovalen dalam hal jenis dan jumlah, antara lain, secara kolektif dikenal sebagai kode histone. Modifikasi ini meliputi fosforilasi, metilasi, asetilasi, ubiquitinasi, dan sumoylasi residu arginin dan lisin pada N-termini histon.
Setiap perubahan, dalam hubungannya dengan yang lain dalam molekul yang sama atau dalam residu histon lain, terutama histon H3, akan menentukan ekspresi atau tidak dari DNA terkait, serta tingkat pemadatan kromatin.
Sebagai aturan umum, telah terlihat, misalnya, bahwa histon hipermetilasi dan hipoasetilasi menentukan bahwa DNA terkait tidak diekspresikan dan bahwa kromatin hadir dalam keadaan yang lebih kompak (heterokromatik, dan karenanya tidak aktif).
Sebaliknya, DNA eukromatik (kurang kompak, dan aktif secara genetik) dikaitkan dengan kromatin yang histonnya mengalami hiperasetilasi dan hipometilasi.
Eukromatin vs heterokromatin
Kita telah melihat bahwa status modifikasi kovalen dari histon dapat menentukan tingkat ekspresi dan pemadatan kromatin lokal. Pada tingkat global, pemadatan kromatin juga diatur oleh modifikasi kovalen histon dalam nukleosom.
Telah ditunjukkan, misalnya, bahwa heterokromatin konstitutif (yang tidak pernah diekspresikan, dan tersusun rapat) cenderung melekat pada lamina nukleus, meninggalkan pori-pori nukleus bebas.
Untuk bagiannya, eukromatin konstitutif (yang selalu diekspresikan, seperti yang mencakup gen pemeliharaan sel, dan terletak di daerah kromatin yang longgar), melakukannya dalam lingkaran besar yang memaparkan DNA untuk ditranskripsi ke mesin transkripsi .
Daerah lain dari DNA genom berosilasi antara dua keadaan ini tergantung pada waktu perkembangan organisme, kondisi pertumbuhan, identitas sel, dll.
Fungsi lainnya
Untuk memenuhi rencana mereka untuk pengembangan, ekspresi dan pemeliharaan sel, genom organisme eukariotik harus mengatur dengan baik kapan dan bagaimana potensi genetik mereka harus terwujud.
Mulai kemudian dari informasi yang disimpan dalam gen mereka, ini terletak di nukleus di daerah tertentu yang menentukan keadaan transkripsi mereka.
Oleh karena itu, kita dapat mengatakan bahwa peran mendasar lainnya dari nukleosom, melalui perubahan kromatin yang membantu untuk mendefinisikannya, adalah organisasi atau arsitektur nukleus yang menampungnya.
Arsitektur ini diwarisi dan dilestarikan secara filogenetik berkat keberadaan unsur modular dari kemasan informasi ini.
Referensi
- Alberts, B., Johnson, AD, Lewis, J., Morgan, D., Raff, M., Roberts, K., Walter, P. (2014) Biologi Molekuler Sel ( Edisi ke- 6 ). WW Norton & Company, New York, NY, AS.
- Brooker, RJ (2017). Genetika: Analisis dan Prinsip. Pendidikan Tinggi McGraw-Hill, New York, NY, AS.
- Cosgrove, MS, Boeke, JD, Wolberger, C. (2004). Mobilitas nukleosom yang diatur dan kode histon. Biologi Struktural & Molekuler Alam, 11: 1037-43.
- Cukup baik, UW (1984) Genetika. WB Saunders Co. Ltd, Pkiladelphia, PA, AS.
- Griffiths, AJF, Wessler, R., Carroll, SB, Doebley, J. (2015). Sebuah Pengantar Analisis Genetika (11 th ed.). New York: WH Freeman, New York, NY, AS.
