teori membran lipat mengusulkan bahwa membran organel berasal dari ekspansi dan invaginasi dari membran plasma. JD Robertson, pelopor dalam mikroskop elektron, mencatat pada tahun 1962 bahwa beberapa badan intraseluler memiliki struktur yang secara visual identik dengan membran plasma.
Ide struktur yang membatasi sel muncul segera setelah konsep “sel” muncul, sehingga beberapa penelitian dilakukan untuk menjelaskan karakteristik struktur ini.
Membran plasma
Indeks artikel
membran plasma
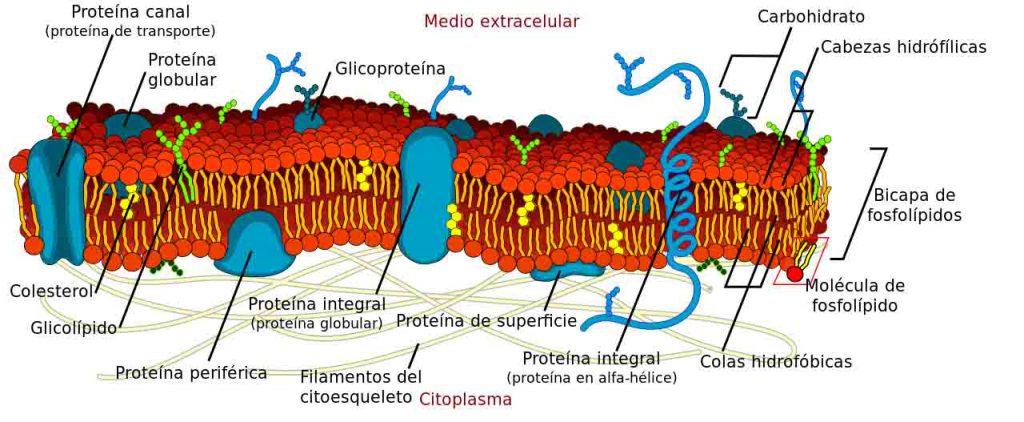
Membran plasma adalah struktur yang dibentuk oleh lapisan ganda fosfolipid yang diatur sedemikian rupa sehingga kelompok polar berorientasi ke sitosol dan media ekstraseluler, sedangkan kelompok apolar diatur ke arah bagian dalam membran.
Fungsi utamanya adalah untuk mendefinisikan sel, baik eukariotik maupun prokariotik, karena secara fisik memisahkan sitoplasma dari lingkungan ekstraseluler.
Terlepas dari fungsi strukturalnya, diketahui bahwa membran tidak statis, melainkan penghalang elastis dan dinamis di mana sejumlah besar proses penting untuk sel berlangsung.
Beberapa proses yang terjadi di membran adalah penahan sitoskeletal, transportasi molekul, pensinyalan, dan koneksi dengan sel lain untuk membentuk jaringan. Selain itu, berbagai macam organel juga memiliki membran di mana proses lain yang sangat penting terjadi.
Latar belakang teori pelipatan membran
Studi elektrofisiologi
Jauh sebelum Robertson mengajukan teori pelipatan membran pada tahun 1962, penelitian pertama kali dilakukan untuk menentukan seperti apa struktur ini. Dengan tidak adanya mikroskop elektron, studi elektrofisiologis mendominasi, di antaranya yang berikut ini menonjol:
1895
Overton mencatat bahwa lipid melintasi membran sel lebih mudah daripada molekul alam lain, oleh karena itu, ia menyimpulkan bahwa membran harus terdiri, sebagian besar, lipid.
1902
J. Bernstein mempresentasikan hipotesisnya, yang menyebutkan bahwa sel terdiri dari larutan dengan ion bebas yang dibatasi oleh lapisan tipis yang tidak dapat ditembus oleh molekul bermuatan tersebut.
1923
Fricke mengukur kemampuan membran eritrosit untuk menyimpan muatan (kapasitansi), menentukan bahwa nilai ini adalah 0,81 F / cm 2 .
Kemudian ditentukan bahwa membran jenis sel lain memiliki nilai kapasitansi yang sama, oleh karena itu, membran harus menjadi struktur kesatuan.
1925
Gorter dan Grendel mengukur luas eritrosit mamalia dengan bantuan mikroskop. Mereka kemudian mengekstrak lipid dari sejumlah jenis sel yang diketahui dan mengukur area yang mereka tempati.
Mereka memperoleh rasio 1:2 sel: membran sebagai hasilnya. Ini berarti bahwa membran sel adalah struktur ganda, sehingga menimbulkan istilah “lapisan ganda lipid”.
1935
Studi sebelum 1935 menyarankan adanya protein dalam membran, hal ini menyebabkan Danielli dan Davson mengusulkan caral Sandwich atau caral Protein-Lipid-Protein.
Menurut caral ini, membran plasma terdiri dari dua lapisan fosfolipid yang ditemukan di antara dua lapisan protein, yang terkait dengan membran melalui interaksi elektrostatik.
Studi mikroskop elektron
Pada tahun 1959, berkat munculnya mikroskop elektron, J. David Robertson mengumpulkan cukup bukti untuk mengkonfirmasi dan melengkapi caral yang diusulkan oleh Gorter dan Grendel (1925) dan Danielli dan Davson (1935), dan untuk mengusulkan caral “Membran Kesatuan”.
Model ini mempertahankan karakteristik caral yang diusulkan oleh Danielli dan Davson dari lipid bilayer, dengan variasi lapisan protein yang dalam hal ini asimetris dan terputus-putus.
Apa yang dimaksud dengan teori lipatan membran?
Kedatangan mikroskop elektron memungkinkan kita untuk memiliki gagasan yang cukup jelas tentang bagaimana membran plasma terbentuk.
Namun, fakta ini disertai dengan visualisasi beberapa membran intracytoplasmic yang membentuk kompartemen intraseluler, yang membuat Robertson pada tahun 1962 mengusulkan “Teori Lipatan Membran”.
Teori pelipatan membran adalah bahwa membran plasma meningkat permukaannya dan berinvaginasi untuk menimbulkan membran intracytoplasmic, membran ini mengelilingi molekul yang berada di sitosol, sehingga berasal dari organel.
Menurut teori ini, selubung nukleus, retikulum endoplasma , aparatus Golgi , lisosom , dan vakuola dapat berasal dari cara ini.
Kontinuitas yang ada antara membran plasma dengan tiga organel pertama yang disebutkan di atas telah dikonfirmasi oleh studi mikroskop elektron pada jenis sel yang berbeda.
Namun, Robertson juga mengusulkan dalam teorinya bahwa organel vesikular seperti lisosom dan vakuola juga berasal dari invaginasi yang kemudian terpisah dari membran.
Karena karakteristik dari teori pelipatan membran, itu dianggap sebagai perpanjangan dari caral membran unit yang dia usulkan sendiri pada tahun 1959.
Mikrograf yang diambil oleh Robertson menunjukkan bahwa semua membran ini sama dan oleh karena itu mereka harus memiliki komposisi yang cukup mirip.
Namun, spesialisasi organel sangat mengubah komposisi membran, yang mengurangi sifat yang sama pada tingkat biokimia dan molekuler.
Dengan cara yang sama, fakta bahwa membran memiliki fungsi utamanya adalah berfungsi sebagai penghalang stabil untuk media berair dipertahankan.
Pentingnya teori ini
Berkat semua tes yang dilakukan antara tahun 1895 dan 1965, terutama studi mikroskopis yang dilakukan oleh JD Robertson, pentingnya membran sel ditekankan.
Dari caral kesatuannya, peran penting yang dimainkan membran dalam struktur dan fungsi sel mulai disorot, sedemikian rupa sehingga studi tentang struktur ini dianggap sebagai masalah mendasar dalam biologi saat ini.
Sekarang, sehubungan dengan kontribusi teori pelipatan membran, ini saat ini tidak diterima. Namun, pada saat itu menyebabkan lebih banyak ahli di bidang itu mencoba menjelaskan asal tidak hanya membran sel, tetapi juga asal sel eukariotik itu sendiri, seperti yang dilakukan Lynn Margulis pada tahun 1967 ketika mengangkat teori endosimbiotik.
Referensi
- Lodish H, Berk A, Kaiser C, dkk. Biologi Sel Molekuler . Vol 39. Edisi ke-8.; 2008.
- Heuser JE. Dalam Memori J. David Robertson. Am Soc Cell Biol . 1995: 11-13. Diambil dari heuserlab.wustl.edu.
- Lee A. Struktur membran. Curr Biol . 2001; 11 (20): R811-R814. doi: 10.1083 / jcb.91.3.189s.
- Gupta GP. Biologi Sel Tumbuhan . Rumah Penerbitan Penemuan; 2004. Diambil dari books.google.
- Membran Heimburg T. — Sebuah Pengantar. Term Biophys Anggota . 2007; (2001): 1-13. doi: 10.1002 / 9783527611591.ch1.
- Stoeckenius W, Stoeckenius W, Engelman DM, Engelman DM. Tinjau caral terkini untuk struktur membran biologis. J Sel Biol .