
sintesis lipid terdiri dari serangkaian reaksi enzimatik dengan cara yang hidrokarbon rantai pendek kental untuk membentuk molekul rantai panjang yang kemudian dapat mengalami modifikasi kimia yang berbeda.
Lipid adalah kelas biomolekul yang sangat bervariasi yang disintesis oleh semua sel hidup dan yang terspesialisasi dalam berbagai fungsi penting untuk pemeliharaan kehidupan seluler.
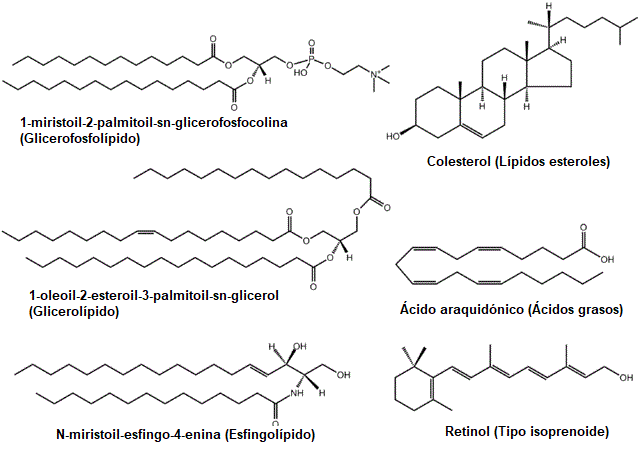
Beberapa contoh lipid umum: gliserofosfolipid, sterol, gliserolipid, asam lemak, sphingolipid, dan prenol (Sumber: Pengunggah asli adalah Lmaps di Wikipedia bahasa Inggris. / GFDL 1.2 (http://www.gnu.org/licenses/old-licenses/ fdl-1.2.html) melalui Commons, diadaptasi oleh Raquel Parada)
Lipid adalah komponen utama membran biologis, fakta yang menjadikannya molekul mendasar bagi keberadaan sel sebagai entitas yang terisolasi dari lingkungannya.
Beberapa lipid juga memiliki fungsi khusus seperti pigmen, kofaktor, transporter, deterjen, hormon, pembawa pesan intra dan ekstraseluler, jangkar kovalen untuk protein membran, dll. Oleh karena itu, kemampuan untuk mensintesis berbagai jenis lipid sangat penting untuk kelangsungan hidup semua organisme hidup.
Kelompok besar senyawa ini secara tradisional diklasifikasikan ke dalam beberapa kategori atau subkelompok: asam lemak (jenuh dan tidak jenuh), gliserida (fosfogliserida dan gliserida netral), lipid non-gliserida (sphingolipids (sphingomyelins dan glikolipid), steroid dan lilin) dan lipid kompleks ( lipoprotein).
Indeks artikel
Jenis lipid dan mekanisme sintesis utamanya
Semua urutan reaksi dari jalur biosintesis lipid adalah endergonik dan reduktif. Dengan kata lain, mereka semua menggunakan ATP sebagai sumber energi dan pembawa elektron tereduksi, seperti NADPH, sebagai daya pereduksi.
Selanjutnya, reaksi utama dari jalur biosintetik jenis utama lipid akan dijelaskan, yaitu asam lemak dan eikosanoid, triasilgliserol dan fosfolipid dan sterol (kolesterol).
– Sintesis asam lemak
Asam lemak adalah molekul yang sangat penting dari sudut pandang lipid, karena merupakan bagian dari lipid yang paling relevan dalam sel. Sintesisnya, bertentangan dengan apa yang dipikirkan banyak ilmuwan selama studi pertama dalam hal ini, tidak terdiri dari rute kebalikan dari -oksidasinya.
Faktanya, jalur metabolisme ini terjadi di kompartemen sel yang berbeda dan membutuhkan partisipasi dari zat antara tiga karbon yang dikenal sebagai malonil-KoA, yang tidak diperlukan untuk oksidasi.
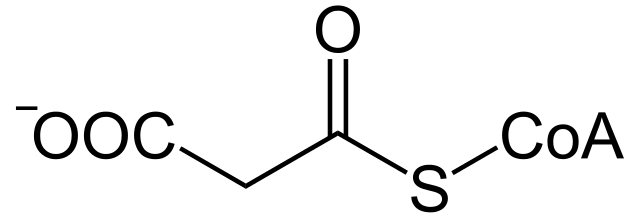
Malonil-KoA. NEUROtiker / Domain publik
Lebih jauh lagi, ini terkait erat dengan kelompok sulfhidril protein yang dikenal sebagai Protein Pembawa Asil (ACP ).
Secara umum, sintesis asam lemak, terutama asam lemak rantai panjang, adalah proses berurutan di mana empat langkah diulang di setiap “putaran”, dan selama setiap putaran dihasilkan gugus asil jenuh yang merupakan substrat untuk berikutnya , yang melibatkan kondensasi lain dengan molekul malonil-KoA baru.
Dalam setiap putaran atau siklus reaksi rantai asam lemak memanjang dua karbon, hingga mencapai panjang 16 atom (palmitat), setelah itu meninggalkan siklus.
Pembentukan malonil-KoA
Zat antara tiga atom karbon ini terbentuk secara ireversibel dari asetil-KoA berkat aksi enzim asetil-KoA karboksilase, yang memiliki gugus prostetik biotin yang terikat secara kovalen dengan enzim dan berpartisipasi dalam katalisis ini dalam Dua langkah.
Dalam reaksi ini, gugus karboksil yang berasal dari molekul bikarbonat (HCO3-) ditransfer ke biotin dengan cara yang bergantung pada ATP, di mana gugus biotinil bertindak sebagai “pengangkut sementara” untuk molekul saat mentransfernya ke asetil-Koa. , menghasilkan malonil-KoA.
Dalam urutan sintesis asam lemak, reduktor yang digunakan adalah NADPH dan gugus pengaktifnya adalah dua gugus tiol (-SH) yang merupakan bagian dari kompleks multi-enzim yang disebut sintase asam lemak, yang paling penting dalam katalisis sintetik.
Pada vertebrata , kompleks sintase asam lemak adalah bagian dari rantai polipeptida besar tunggal, di mana 7 aktivitas enzimatik karakteristik dari rute sintesis diwakili, serta aktivitas hidrolitik yang diperlukan untuk melepaskan zat antara pada akhir proses. .

Struktur enzim sintase asam lemak (Sumber: Boehringer Ingelheim / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0) melalui Wikimedia Commons)
7 aktivitas enzimatik kompleks ini adalah: protein pengangkut gugus asil (ACP), asetil-KoA-ACP transasetilase (AT), -ketoasil-ACP sintase (KS), malonil-KoA-ACP transferase (MT), – ketoasil -ACP reduktase (KR), -hidroksiasil-ACP dehidratase (HD) dan enoyl-ACP reduktase (ER).
Sebelum reaksi kondensasi dapat terjadi untuk merakit rantai asam lemak, dua gugus tiol dalam kompleks enzim menjadi “bermuatan” dengan gugus asil: pertama, asetil-KoA ditransfer ke gugus -SH dari sistein di -ketoasil-ACP sintase bagian dari kompleks, suatu reaksi yang dikatalisis oleh enzim asetil-KoA-ACP transasetilase (AT).
Selanjutnya, gugus malonil ditransfer dari molekul malonil-KoA ke gugus -SH dari bagian pembawa gugus asil (ACP) dari kompleks enzim, suatu reaksi yang dikatalisis oleh enzim malonil-KoA-ACP transferase (MT), yang juga Ini adalah bagian dari kompleks sintase asam lemak.
Urutan empat reaksi untuk setiap “putaran” siklus reaksi adalah sebagai berikut:
- Kondensasi: Gugus asetil dan malonil “bermuatan” pada enzim mengembun untuk membentuk molekul asetoasetil-ACP, yang melekat pada bagian ACP melalui gugus -SH. Pada langkah ini, molekul CO2 diproduksi dan dikatalisis oleh -ketoasil-ACP sintase (gugus asetil menempati posisi “metil terminal” dari kompleks asetoasetil-ACP).
- Reduksi gugus karbonil: gugus karbonil pada posisi C3 asetoasetil-ACP direduksi menjadi D-β-hidroksibutiril-ACP, suatu reaksi yang dikatalisis oleh -ketoasil-ACP reduktase, yang menggunakan NADPH sebagai donor elektron.
- Dehidrasi: karbon C2 dan C3 dari D-β-hydroxybutyryl-ACP tidak memiliki molekul air, membentuk ikatan rangkap yang berakhir dengan produksi senyawa baru trans -∆2-butenoyl-ACP. Proses ini dimediasi oleh enzim -hidroksiasil-ACP dehidratase (HD).
- Reduksi ikatan rangkap: ikatan rangkap senyawa yang terbentuk pada tahap dehidrasi dijenuhkan (direduksi) menghasilkan butiril-ACP melalui reaksi yang dikatalisis oleh enzim enoil-ACP reduktase (ER), yang juga menggunakan NADPH sebagai zat pereduksi.
Reaksi sintesis terjadi sampai terbentuk molekul palmitat (16 atom karbon), yang dihidrolisis dari kompleks enzimatik dan dilepaskan sebagai kemungkinan prekursor untuk asam lemak dengan rantai lebih panjang, yang dihasilkan oleh sistem pemanjangan. bagian dari retikulum endoplasma dan di mitokondria.
Modifikasi lain yang dapat dialami oleh molekul-molekul ini, seperti desaturasi, misalnya, dikatalisis oleh enzim yang berbeda, yang umumnya terjadi di retikulum endoplasma halus.
– Sintesis eicosanoids
Eicosanoids adalah lipid seluler yang berfungsi sebagai molekul pembawa pesan “jarak pendek”, yang diproduksi oleh beberapa jaringan untuk berkomunikasi dengan sel-sel di jaringan tetangganya. Molekul-molekul ini disintesis dari asam lemak tak jenuh ganda dari 20 atom karbon.
Prostaglandin
Sebagai respons terhadap stimulus hormonal, enzim fosfolipase A menyerang fosfolipid membran dan melepaskan arakidonat dari 2-karbon gliserol. Senyawa ini diubah menjadi prostaglandin berkat enzim retikulum endoplasma halus dengan aktivitas bifungsional: siklooksigenase (COX) atau prostaglandin H2 sintase.
Tromboksan
Prostaglandin dapat diubah menjadi tromboksan berkat adanya tromboksan sintase dalam trombosit darah (trombosit). Molekul-molekul ini terlibat dalam langkah awal pembekuan darah.
– Sintesis triasilgliserol
Asam lemak adalah molekul penting untuk sintesis senyawa lain yang lebih kompleks dalam sel, seperti triasilgliserol atau lipid membran gliserofosfolipid (proses yang bergantung pada kebutuhan metabolisme seluler).
Hewan menghasilkan triasilgliserol dan gliserofosfolipid dari dua prekursor umum: asil-KoA lemak dan L-gliserol 3-fosfat. Asil-KoA lemak diproduksi oleh enzim asil-KoA sintetase yang berpartisipasi dalam -oksidasi, sedangkan L-gliserol 3-fosfat diperoleh dari glikolisis dan oleh aksi dua enzim alternatif: gliserol 3-fosfat dehidrogenase dan gliserol kinase.
Triasilgliserol dibentuk oleh reaksi antara dua molekul asil-KoA lemak dan molekul diasilgliserol 3-fosfat; Reaksi transfer ini dikatalisis oleh asil transferase spesifik.
Dalam reaksi ini, asam fosfatidat awalnya diproduksi, yang didefosforilasi oleh enzim asam fosfatidat fosfatase untuk menghasilkan 1,2-diasilgliserol, yang lagi-lagi mampu menerima molekul ketiga asil-KoA lemak, menghasilkan triasilgliserol.
– Sintesis fosfolipid
Fosfolipid adalah molekul yang sangat bervariasi, karena banyak yang berbeda dapat dibentuk oleh kombinasi asam lemak dan kelompok “kepala” yang berbeda dengan tulang punggung gliserol (gliserofosfolipid) atau sphingosine (sphingolipids) yang menjadi ciri mereka.
Perakitan umum molekul-molekul ini membutuhkan sintesis tulang punggung gliserol atau sfingosin, penyatuan dengan asam lemak yang sesuai, baik dengan esterifikasi atau amidasi, penambahan gugus “kepala” hidrofilik melalui ikatan fosfodiester dan, jika perlu, perubahan atau pertukaran kelompok terakhir ini.
Pada eukariota proses ini terjadi di retikulum endoplasma halus dan juga di membran mitokondria bagian dalam, di mana mereka dapat tetap tidak terbatas atau dari mana mereka dapat dipindahkan ke tempat lain.
Langkah-langkah reaksi
Langkah pertama dari reaksi sintesis gliserofosfolipid setara dengan produksi triasilgliserol, karena molekul gliserol 3-fosfat diesterifikasi menjadi dua molekul asam lemak pada karbon 1 dan 2, membentuk asam fosfatidat. Adalah umum untuk menemukan fosfolipid yang memiliki asam lemak jenuh dalam C1 dan tak jenuh dalam C2 gliserol.
Asam fosfatidat juga dapat diproduksi dengan fosforilasi molekul diasilgliserol yang sudah disintesis atau “daur ulang”.
Gugus “kepala” polar dari molekul ini dibentuk melalui ikatan fosfodiester. Hal pertama yang harus terjadi agar proses ini terjadi dengan benar adalah “aktivasi” salah satu gugus hidroksil yang berpartisipasi dalam proses dengan mengikat nukleotida seperti cytidine diphosphate (CDP), yang secara nukleofilik digantikan oleh gugus lain. hidroksil yang berpartisipasi dalam reaksi.
Jika molekul ini berikatan dengan diasilgliserol, maka CDP-diasilgliserol (bentuk asam fosfatidat yang “diaktifkan”) akan terbentuk, tetapi ini juga dapat terjadi pada gugus hidroksil dari gugus “kepala”.
Dalam kasus fosfatidilserin, misalnya, diasilgliserol diaktifkan oleh kondensasi molekul asam fosfatidat dengan molekul sitidin trifosfat (CTP), membentuk CDP-diasilgliserol dan menghilangkan pirofosfat.
Jika molekul CMP (cytidine monophosphate) digantikan oleh serangan nukleofilik hidroksil serin atau hidroksil pada 1-karbon gliserol 3-fosfat, fosfatidilserin atau fosfatidilgliserol 3-fosfat dapat dilepaskan, dari mana monoester fosfat dapat dilepaskan. dapat dilepaskan dan menghasilkan fosfatidilgliserol.
Kedua molekul yang dihasilkan dengan cara ini berfungsi sebagai prekursor untuk lipid membran lainnya, yang sering berbagi jalur biosintetik satu sama lain.
– Sintesis kolesterol
Kolesterol merupakan molekul esensial bagi hewan yang dapat disintesis oleh selnya, sehingga tidak esensial dalam makanan sehari-hari. Molekul 27 atom karbon ini dihasilkan dari prekursor: asetat.
Molekul kompleks ini terbentuk dari asetil-KoA dalam empat tahap utama:
- Kondensasi tiga unit asetat untuk membentuk mevalonat, molekul antara 6-karbon (pertama molekul asetoasetil-KoA dibentuk dengan dua asetil-KoA (enzim tiolase) dan kemudian satu lagi dari -hidroksi-β-metilglutaril-KoA ( HMG- CoA) (enzim sintetase HMG-CoA) Mevalonat terbentuk dari HMG-CoA dan berkat enzim HMG-CoA reduktase.
- Konversi unit mevalonat menjadi isoprena. 3 gugus fosfat pertama ditransfer dari 3 molekul ATP ke mevalonat. Salah satu fosfat hilang bersama dengan gugus karbonil yang berdekatan dan 3-isopentenil pirofosfat terbentuk, yang diisomerisasi untuk menghasilkan dimetilalil pirofosfat.
- Polimerisasi atau kondensasi 6 C 5 unit isoprena untuk membentuk C 30 squalene (molekul linier).
- Siklisasi squalene untuk membentuk 4 cincin inti steroid kolesterol dan perubahan kimia berikutnya: oksidasi, migrasi dan eliminasi gugus metil, dll., yang menghasilkan kolesterol.
Referensi
- Garrett, RH, & Grisham, CM (2001). Prinsip-prinsip biokimia: dengan fokus manusia. Perusahaan Penerbitan Brooks / Cole.
- Murray, RK, Granner, DK, Mayes, PA, & Rodwell, VW (2014). Biokimia Harper yang diilustrasikan. Mcgraw-Hill.
- Nelson, DL, Lehninger, AL, & Cox, MM (2008). Prinsip biokimia Lehninger. Macmillan.
- Jacquemyn, J., Cascalho, A., & Goodchild, RE (2017). Seluk-beluk retikulum endoplasma – biosintesis lipid yang dikendalikan. Laporan EMBO, 18 (11), 1905-1921.
- Ohlrogge, J., & Telusuri, J. (1995). Biosintesis lipid. Sel Tumbuhan, 7 (7), 957.
