
phosphatidylethanolamine (PE) adalah kelimpahan gliserofosfolipid dalam membran plasma prokariota . Sebaliknya, dalam membran sel eukariotik, ini adalah gliserofosfolipid paling melimpah kedua di bagian dalam membran plasma setelah fosfatidilkolin.
Meskipun phosphatidylethanolamine berlimpah, kelimpahannya tidak hanya tergantung pada jenis sel tetapi juga pada kompartemen dan momen spesifik dari siklus hidup sel yang dipertimbangkan.
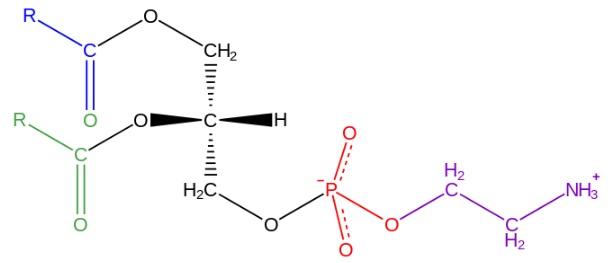
Molekul fosfatidiletanolamin
Membran biologis adalah penghalang yang mendefinisikan organisme seluler. Mereka tidak hanya memiliki fungsi perlindungan dan isolasi, tetapi mereka juga merupakan kunci untuk pembentukan protein yang membutuhkan lingkungan hidrofobik untuk berfungsi optimal.
Baik eukariota dan prokariota memiliki membran yang terutama terdiri dari gliserofosfolipid dan pada tingkat lebih rendah sphingolipid dan sterol.
Gliserofosfolipid adalah molekul amfipatik yang terstruktur pada tulang punggung L-gliserol yang diesterifikasi pada posisi sn-1 dan sn-2 oleh dua asam lemak dengan panjang dan derajat kejenuhan yang bervariasi. Dalam hidroksil dari posisi sn-3 itu diesterifikasi oleh gugus fosfat, yang pada gilirannya berbagai jenis molekul dapat dilampirkan yang menimbulkan berbagai kelas gliserofosfolipid.
Di dunia seluler terdapat berbagai macam gliserofosfolipid, namun yang paling melimpah adalah fosfatidilkolin (PC), fosfatidiletanolamin (PE), fosfatidilserin (PS), fosfatidilinositol (PI), asam fosfatidat (PA), fosfatidilgliserol (PG) dan kardiolipin. (CL).
Indeks artikel
Struktur
Struktur phosphatidylethanolamine ditemukan oleh Baer et al.Pada tahun 1952. Seperti yang telah ditentukan secara eksperimental untuk semua gliserofosfolipid, phosphatidylethanolamine terdiri dari molekul gliserol yang diesterifikasi pada posisi sn-1 dan sn-2 dengan rantai asam Lemak antara 16 dan 20 atom karbon.
Asam lemak yang teresterifikasi pada hidroksil sn-1 umumnya jenuh (tanpa ikatan rangkap) dengan panjang maksimum 18 atom karbon, sedangkan rantai yang terikat pada posisi sn-2 lebih panjang dan dengan satu atau lebih tidak jenuh (ikatan rangkap).
Derajat kejenuhan rantai ini berkontribusi pada elastisitas membran, yang memiliki pengaruh besar pada penyisipan dan penyerapan protein di bilayer.
phosphatidylethanolamine dianggap nonlamellar gliserofosfolipid dan memiliki geometri kerucut. Bentuk ini diberikan oleh ukuran kecil dari kelompok kutub atau “kepala”, dalam kaitannya dengan rantai asam lemak yang terdiri dari “ekor” hidrofobik.
Gugus “kepala” atau polar dari fosfatidiletanolamin memiliki sifat zwitterionik, yaitu memiliki gugus yang dapat bermuatan positif dan negatif pada kondisi pH tertentu.
Karakteristik ini memungkinkan untuk ikatan hidrogen dengan sejumlah besar residu asam amino, dan distribusi muatannya merupakan penentu penting untuk topologi domain banyak protein membran integral.
Biosintesis
Dalam sel eukariotik, sintesis lipid struktural dibatasi secara geografis, situs biosintesis utama adalah retikulum endoplasma (ER) dan pada tingkat lebih rendah aparatus Golgi .
Ada empat jalur biosintetik independen untuk produksi phosphatidylethanolamine: (1) jalur CDP-ethanolamine, juga dikenal sebagai jalur Kennedy; (2) jalur PSD untuk dekarboksilasi fosfatidilserin (PS); (3) asilasi liso-PE dan (4) reaksi perubahan basa dari gugus polar gliserofosfolipid lain.
Rute Kennedy
Biosintesis fosfatidiletanolamin melalui jalur ini terbatas pada RE dan telah ditunjukkan bahwa dalam sel hati hamster jalur ini merupakan jalur produksi utama. Ini terdiri dari tiga langkah enzimatik berturut-turut dikatalisis oleh tiga enzim yang berbeda.
Pada langkah pertama, fosfoetanolamina dan ADP diproduksi berkat aksi etanolamin kinase, yang mengkatalisis fosforilasi etanolamin yang bergantung pada ATP .
Tidak seperti tumbuhan , baik mamalia maupun ragi tidak mampu memproduksi substrat ini, jadi substrat ini harus dikonsumsi dalam makanan atau diperoleh dari degradasi molekul phosphatidylethanolamine atau sphingosine yang sudah ada sebelumnya.
Phosphoethanolamine digunakan oleh CTP: phosphoethanolamine cytidyltransferase (ET) untuk membentuk senyawa energi tinggi CDP: ethanolamine dan fosfat anorganik.
1,2-Diacylglycerol ethanolamine phosphotransferase (ETP) menggunakan energi yang terkandung dalam ikatan CDP-ethanolamine untuk secara kovalen mengikat ethanolamine ke molekul diasilgliserol yang dimasukkan ke dalam membran, sehingga menghasilkan phosphatidylethanolamine.
Rute PSD
Rute ini beroperasi baik di prokariota dan ragi dan mamalia. Pada bakteri terjadi di membran plasma, tetapi pada eukariota terjadi di area retikulum endoplasma yang terkait erat dengan membran mitokondria.
Pada mamalia jalur ini dikatalisis oleh enzim tunggal, fosfatidilserin dekarboksilase (PSD1p), yang tertanam dalam membran mitokondria, yang gennya dikodekan oleh nukleus. Reaksi melibatkan dekarboksilasi PS menjadi fosfatidiletanolamin.
Dua jalur yang tersisa (asilasi PE-liso dan pertukaran kalsium yang bergantung pada kelompok kutub) terjadi di retikulum endoplasma, tetapi tidak berkontribusi secara signifikan terhadap total produksi fosfatidiletanolamin dalam sel eukariotik.
Fitur
Gliserofosfolipid memiliki tiga fungsi utama di dalam sel, di antaranya fungsi struktural, penyimpanan energi, dan pensinyalan sel yang menonjol.
phosphatidylethanolamine dikaitkan dengan penahan, stabilisasi dan pelipatan beberapa protein membran, serta perubahan konformasi yang diperlukan untuk berfungsinya banyak enzim.
Ada bukti eksperimental yang mengusulkan fosfatidiletanolamin sebagai gliserofosfolipid penting pada tahap akhir telofase, selama pembentukan cincin kontraktil dan pembentukan fragmoplast yang memungkinkan pembelahan membran dua sel anak.
Ini juga memiliki peran penting dalam semua proses fusi dan fisi (penyatuan dan pemisahan) membran retikulum endoplasma dan aparatus Golgi.
Dalam E. coli telah ditunjukkan bahwa phosphatidylethanolamine diperlukan untuk pelipatan yang benar dan fungsi enzim laktosa permease, itulah sebabnya telah disarankan bahwa ia memainkan peran sebagai “pendamping” molekuler.
phosphatidylethanolamine adalah ethanolamine molekul donor utama yang diperlukan untuk modifikasi pasca-translasi banyak protein, seperti jangkar GPI.
Gliserofosfolipid ini merupakan prekursor banyak molekul dengan aktivitas enzimatik. Selain itu, molekul yang berasal dari metabolismenya, serta diasilgliserol, asam fosfatidat, dan beberapa asam lemak, dapat bertindak sebagai pembawa pesan kedua. Selain itu, ini adalah substrat penting untuk produksi fosfatidilkolin.
Referensi
- Brouwers, JFHM, Vernooij, EAAM, Tielens, AGM, & van Golde, LMG (1999). Pemisahan cepat dan identifikasi spesies molekul phosphatidylethanolamine. Jurnal Penelitian Lipid, 40 (1), 164-169. Dipulihkan dari jlr.org
- Calzada, E., McCaffery, JM, & Claypool, SM (2018). Phosphatidylethanolamine yang diproduksi di membran mitokondria bagian dalam sangat penting untuk fungsi kompleks sitokrom bc1 ragi 3. BioRxiv, 1, 46.
- Calzada, E., Onguka, O., & Claypool, SM (2016). Metabolisme Phosphatidylethanolamine dalam Kesehatan dan Penyakit. Tinjauan Internasional Biologi Sel dan Molekuler (Vol. 321). Elsevier Inc.
- Gibellini, F., & Smith, TK (2010). Sintesis jalur-de novo Kennedy dari fosfatidiletanolamin dan fosfatidilkolin. IUBMB Life, 62 (6), 414–428.
- Harayama, T., & Riezman, H. (2018). Memahami keragaman komposisi lipid membran. Nature Review Biologi Sel Molekuler, 19 (5), 281-296.
- Lucky, M. (2008). Biologi struktural membran: dengan dasar biokimia dan biofisika. Pers Universitas Cambridge. Dipulihkan dari cambrudge.org
- Seddon, JM, Cevc, G., Kaye, RD, & Marsh, D. (1984). Studi Difraksi sinar-X dari Polimorfisme Diasil- dan Dialkylphosphatidylethanolamines terhidrasi. Biokimia, 23 (12), 2634-2644.
- Sendecki, AM, Poyton, MF, Baxter, AJ, Yang, T., & Cremer, PS (2017). Mendukung Lipid Bilayers dengan Phosphatidylethanolamine sebagai Komponen Utama. Langmuir, 33 (46), 13423–13429.
- van Meer, G., Voelker, DR, & Feignenson, GW (2008). Lipid membran: di mana mereka berada dan bagaimana mereka berperilaku. Ulasan Alam, 9, 112-124.
- Vance, JE (2003). Biologi Molekuler dan Sel Metabolisme Phosphatidylserine dan Phosphatidylethanolamine. Dalam K. Moldave (Ed.), Kemajuan Penelitian Asam Nukleat dan Biologi Molekuler (hlm. 69-111). Pers Akademik.
- Vance, JE (2008). Phosphatidylserine dan phosphatidylethanolamine dalam sel mamalia: dua aminofosfolipid yang terkait secara metabolik. Jurnal Penelitian Lipid, 49 (7), 1377–1387.
- Vance, JE, & Tasseva, G. (2013). Pembentukan dan fungsi phosphatidylserine dan phosphatidylethanolamine dalam sel mamalia. Biochimica et Biophysica Acta – Biologi Molekuler dan Sel Lipid, 1831 (3), 543–554.
- Watkins, SM, Zhu, X., & Zeisel, SH (2003). Aktivitas Phosphatidylethanolamine-N-methyltransferase dan diet kolin mengatur fluks lipid plasma hati dan metabolisme asam lemak esensial pada tikus. Jurnal Nutrisi, 133 (11), 3386-3391.
