
glikolisis atau glikolisis adalah rute utama dari katabolisme glukosa, yang tujuan utamanya adalah untuk menghasilkan energi di dalam bentuk ATP dan kekuasaan mengurangi di dalam bentuk NADH, dari karbohidrat ini.
Jalur ini, dijelaskan sepenuhnya pada tahun 1930-an oleh Gustav Embden dan Otto Meyerhof saat mempelajari pengambilan glukosa dalam sel otot rangka, melibatkan oksidasi lengkap monosakarida dan, dengan sendirinya, merupakan jalur anaerobik untuk glukosa, memperoleh energi.
Struktur molekul ATP, salah satu produk glikolitik (Ringkasan jalur glikolitik (Sumber: Tekks di Wikipedia bahasa Inggris / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0) melalui Wikimedia Commons) melalui Wikimedia Commons)
Ini adalah salah satu rute metabolisme utama, karena terjadi, dengan perbedaannya, di semua organisme hidup yang ada, uniseluler atau multiseluler, prokariotik atau eukariotik, dan dianggap sebagai rantai reaksi yang secara evolusioner sangat kekal di alam.
Faktanya, ada beberapa organisme dan jenis sel yang bergantung secara eksklusif pada rute ini untuk bertahan hidup.
Dalam contoh pertama, glikolisis terdiri dari oksidasi glukosa, dari 6 atom karbon, menjadi piruvat, yang memiliki tiga atom karbon; dengan produksi ATP dan NADH secara bersamaan, berguna untuk sel dari sudut pandang metabolisme dan sintetik.
Dalam sel yang mampu memproses lebih lanjut produk yang diperoleh dari katabolisme glukosa, glikolisis berakhir dengan produksi karbon dioksida dan air melalui siklus Krebs dan rantai transpor elektron (glikolisis aerobik).
Sepuluh reaksi enzimatis berlangsung selama jalur glikolitik dan, meskipun pengaturan reaksi ini mungkin agak berbeda dari satu spesies ke spesies lain, mekanisme pengaturannya juga cukup dipertahankan.
Indeks artikel
Fungsi Glikolisis
Dari sudut pandang metabolisme, glukosa adalah salah satu karbohidrat terpenting bagi semua makhluk hidup.
Ini adalah molekul yang stabil dan sangat larut, sehingga dapat diangkut dengan relatif mudah ke seluruh tubuh hewan atau tumbuhan, dari mana ia disimpan dan / atau diperoleh ke tempat yang dibutuhkan sebagai bahan bakar seluler.
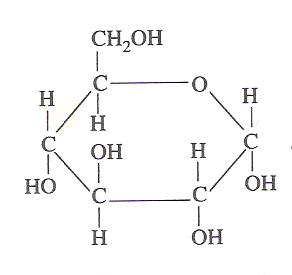
Struktur glukosa (Sumber: Oliva93 / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0) melalui Wikimedia Commons)
Energi kimia yang terkandung dalam glukosa dimanfaatkan oleh sel-sel hidup melalui glikolisis, yang terdiri dari serangkaian langkah yang sangat terkontrol dimana energi yang dilepaskan dari oksidasi karbohidrat ini dapat “ditangkap” menjadi bentuk energi yang lebih bermanfaat. , oleh karena itu pentingnya energi ini. .
Melalui rute ini, tidak hanya energi (ATP) dan daya pereduksi (NADH) yang diperoleh, tetapi juga menyediakan serangkaian perantara metabolisme yang merupakan bagian dari rute lain, juga penting dari fungsi seluler anabolik (biosintetik) dan umum. Berikut daftarnya:
– Glukosa 6-fosfat ke jalur pentosa fosfat (PPP, dari Jalur Pentosa Fosfat Inggris )
– Piruvat untuk fermentasi laktat
– Piruvat untuk sintesis asam amino (terutama alanin)
– Piruvat untuk siklus asam trikarboksilat
– Fruktosa 6-fosfat, glukosa 6-fosfat dan dihidroksiaseton fosfat, yang berfungsi sebagai “bahan penyusun” di jalur lain seperti sintesis glikogen, asam lemak, trigliserida, nukleotida, asam amino, dll.
Produksi energi
Jumlah ATP yang dihasilkan oleh jalur glikolitik, ketika sel yang memproduksinya tidak dapat hidup dalam kondisi aerobik, cukup untuk memasok kebutuhan energi sel ketika digabungkan dengan berbagai jenis proses fermentasi.
Namun, ketika menyangkut sel aerobik, glikolisis juga berfungsi sebagai sumber energi darurat dan berfungsi sebagai “langkah persiapan” sebelum reaksi fosforilasi oksidatif yang menjadi ciri sel dengan metabolisme aerobik.
Enzim yang terlibat dalam glikolisis
Glikolisis hanya dimungkinkan berkat partisipasi 10 enzim yang mengkatalisis reaksi yang menjadi ciri jalur ini. Banyak dari enzim ini bersifat alosterik dan berubah bentuk atau konformasi ketika mereka menjalankan fungsi katalitiknya.
Ada enzim yang memutuskan dan membentuk ikatan kovalen antara substratnya dan ada enzim lain yang memerlukan kofaktor spesifik untuk menjalankan fungsinya, terutama ion logam.
Secara struktural, semua enzim glikolitik memiliki pusat yang pada dasarnya terdiri dari lembaran paralel yang dikelilingi oleh -heliks dan diatur di lebih dari satu domain. Selanjutnya, enzim-enzim ini dicirikan bahwa situs aktifnya biasanya berada di situs pengikatan antara domain.
Penting juga untuk dicatat bahwa regulasi utama jalur melewati kontrol (hormonal atau oleh metabolit) enzim seperti heksokinase, fosfofruktokinase, gliseraldehida 3-fosfat dehidrogenase dan piruvat kinase.
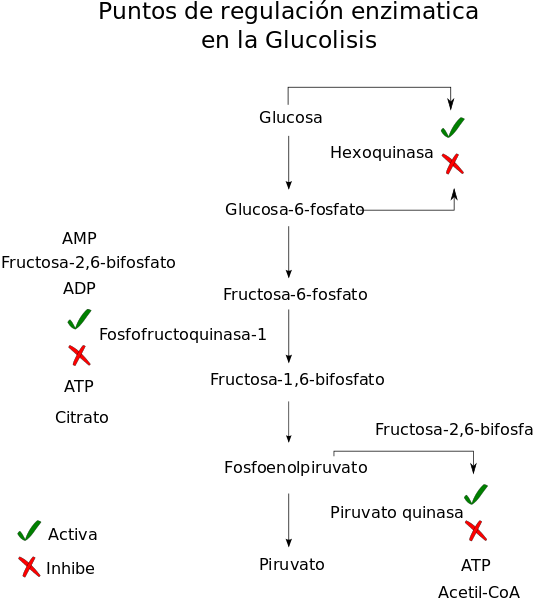
Poin utama regulasi jalur glikolitik (Sumber: Gregor 0492 / CC BY-SA (https://creativecommons.org/licenses/by-sa/3.0) melalui Wikimedia Commons)
1- Heksokinase (HK)
Reaksi glikolisis pertama (fosforilasi glukosa) dikatalisis oleh heksokinase (HK), yang mekanisme kerjanya tampaknya terdiri dari substrat “pengencangan yang diinduksi” yang mempromosikan “penguncian” enzim di sekitar ATP dan glukosa (substratnya) setelah itu telah terikat pada mereka.
Tergantung pada organisme yang dipertimbangkan, mungkin ada satu atau lebih isoenzim, yang berat molekulnya berkisar antara 50 (sekitar 500 asam amino) dan 100 kDa, karena mereka tampaknya mengelompok bersama dalam bentuk dimer, yang pembentukannya disukai oleh adanya glukosa, ion magnesium, dan ATP.
Heksokinase memiliki struktur tersier yang terdiri dari lembaran alfa dan beta terbuka, meskipun ada banyak perbedaan struktural dalam enzim ini.
2- fosfoglukosa isomerase (PGI)
Glukosa terfosforilasi oleh heksokinase diisomerisasi menjadi fruktosa 6-fosfat melalui fosfoglukosa isomerase (PGI), juga dikenal sebagai glukosa 6-fosfat isomerase. Enzim, kemudian, tidak menghilangkan atau menambahkan atom, tetapi menyusunnya kembali pada tingkat struktural.
Ini adalah enzim aktif dalam bentuk dimernya (monomer beratnya kurang lebih 66 kDa) dan terlibat tidak hanya dalam glikolisis, tetapi juga dalam glukoneogenesis, dalam sintesis karbohidrat pada tanaman , dll.
3- Fosfofruktokinase (PFK)
Fruktosa 6-fosfat adalah substrat untuk enzim fosfofruktokinase, yang mampu memfosforilasi ulang molekul ini menggunakan ATP sebagai donor gugus fosforil, menghasilkan fruktosa 1,6-bifosfat.
Enzim ini ada pada bakteri dan mamalia sebagai enzim homotetramerik (terdiri dari empat subunit identik masing-masing 33 kDa untuk bakteri dan 85 kDa masing-masing pada mamalia) dan dalam ragi itu adalah oktamer (terdiri dari subunit yang lebih besar, antara 112 dan 118 kDa).
Ini adalah enzim alosterik, yang berarti bahwa ia diatur secara positif atau negatif oleh beberapa produknya (ADP) dan oleh molekul lain seperti ATP dan sitrat.
4- Aldolasse
Juga dikenal sebagai fruktosa 1,6-bifosfat aldolase, aldolase mengkatalisis pemecahan katalitik fruktosa 1,6-bifosfat menjadi dihidroksiaseton fosfat dan gliseraldehida 3-fosfat dan reaksi sebaliknya, yaitu penyatuan kedua gula untuk pembentukan fruktosa 1 ,6-bifosfat.
Dengan kata lain, enzim ini memotong fruktosa 1,6-bifosfat menjadi dua, melepaskan dua senyawa C3 terfosforilasi. Aldolase juga terdiri dari 4 subunit identik, masing-masing dengan situs aktifnya sendiri.
Keberadaan dua kelas (I dan II) dari enzim ini telah ditentukan, yang dibedakan oleh mekanisme reaksi yang dikatalisisnya dan karena beberapa (yang pertama) terjadi pada bakteri dan eukariota “rendah”, dan yang lainnya (yang kedua). ) terdapat pada bakteri, protista, dan metazoa.
Aldolase eukariotik “lebih tinggi” terdiri dari homotetramer dari subunit berat molekul 40 kDa, masing-masing terdiri dari barel yang terdiri dari 8 / lembar.
5- Triosa fosfat isomerase (TIM)
Kedua triosa terfosforilasi dapat saling dikonversi satu sama lain berkat aksi triosa fosfat isomerase, yang memungkinkan kedua gula digunakan selama glikolisis, memastikan penggunaan penuh setiap molekul glukosa yang memasuki jalur tersebut.
Enzim ini telah digambarkan sebagai enzim “sempurna”, karena mengkatalisis reaksi yang dijelaskan sekitar satu triliun kali lebih cepat daripada yang akan terjadi tanpa partisipasi Anda. Situs aktifnya berada di tengah struktur beta-barrel, karakteristik dari banyak enzim glikolitik.
Ini adalah protein dimer, terdiri dari dua subunit identik sekitar 27 kDa, keduanya dengan struktur globular.
6- Gliseraldehida 3-fosfat dehidrogenase (GAPDH)
Gliseraldehida 3-fosfat yang dihasilkan oleh aksi aldolase dan triosa fosfat isomerase berfungsi sebagai substrat untuk GAPDH, yang merupakan enzim homotetramerik (34-38 kDa setiap subunit) yang secara kooperatif mengikat molekul NAD + di masing-masing dari 4 situs aktifnya , serta 2 ion fosfat atau sulfat.
Dalam langkah jalur ini, enzim memungkinkan fosforilasi salah satu substratnya menggunakan fosfat anorganik sebagai donor gugus fosforil, dengan reduksi bersamaan dua molekul NAD+ dan produksi 1,3-bisfosfogliserat.
7- Fosfogliserat kinase (PGK)
Fosfogliserat kinase bertanggung jawab untuk mentransfer salah satu gugus fosfat 1,3-bisfogliserat ke molekul ADP melalui fosforilasi pada tingkat substrat. Enzim ini menggunakan mekanisme yang mirip dengan yang digunakan oleh heksokinase, karena menutup kontak pada substratnya, melindungi mereka dari molekul air yang mengganggu.
Enzim ini, seperti yang lain yang menggunakan dua atau lebih substrat, memiliki situs pengikatan untuk ADP dan satu lagi untuk gula fosfat.
Tidak seperti enzim lain yang dijelaskan, protein ini adalah monomer 44 kDa dengan struktur bilobal, terdiri dari dua domain dengan ukuran yang sama yang dihubungkan oleh “celah” yang sempit.
8- Fosfogliserat mutase
3-fosfogliserat mengalami perubahan dari gugus fosfat menuju karbon 2, di tengah molekul, yang mewakili situs ketidakstabilan strategis yang memfasilitasi transfer gugus berikutnya ke molekul ATP dalam reaksi terakhir jalur tersebut.
Penataan ulang ini dikatalisis oleh enzim fosfogliserat mutase, enzim dimerik untuk manusia dan tetramerik untuk ragi, dengan ukuran subunit mendekati 27 kDa.
9- Enolase
Enolase mengkatalisis dehidrasi 2-fosfogliserat menjadi fosfoenolpiruvat, langkah yang diperlukan untuk pembentukan ATP dalam reaksi berikutnya.
Ini adalah enzim dimer yang terdiri dari dua subunit 45 kDa yang identik. Itu tergantung pada ion magnesium untuk stabilitasnya dan untuk perubahan konformasi yang diperlukan untuk mengikat substratnya. Ini adalah salah satu enzim yang paling banyak diekspresikan dalam sitosol banyak organisme dan melakukan fungsi selain yang glikolitik.
10- Piruvat kinase
Fosforilasi tingkat substrat kedua yang terjadi dalam glikolisis dikatalisis oleh piruvat kinase, yang bertanggung jawab untuk transfer gugus fosforil dari fosfoenolpiruvat ke ADP dan untuk produksi piruvat.
Enzim ini lebih kompleks daripada enzim glikolitik lainnya dan pada mamalia merupakan enzim homotetramerik (57 kDa / subunit). Ada, pada vertebrata , setidaknya 4 isoenzim: L (dalam hati), R (dalam eritrosit), M1 (dalam otot dan otak) dan M2 (jaringan janin dan jaringan dewasa).
Fase glikolisis (langkah demi langkah)
Jalur glikolitik terdiri dari sepuluh langkah berurutan dan dimulai dengan satu molekul glukosa. Selama proses tersebut, molekul glukosa “diaktifkan” atau “disiapkan” dengan penambahan dua fosfat, membalikkan dua molekul ATP.
Kemudian “dipotong” menjadi dua fragmen dan akhirnya dimodifikasi secara kimiawi beberapa kali, mensintesis empat molekul ATP di sepanjang jalan, sehingga perolehan bersih dalam rute sesuai dengan dua molekul ATP.
Dari penjelasan di atas, dapat disimpulkan bahwa rute dibagi menjadi fase “pembalikan” energi, yang mendasar untuk oksidasi lengkap molekul glukosa, dan fase “perolehan” energi lain, di mana energi yang digunakan awalnya diganti dan dua diperoleh Molekul ATP bersih.
– Fase investasi energi
1- Langkah pertama dari jalur glikolitik terdiri dari fosforilasi glukosa yang dimediasi oleh heksokinase (HK), di mana enzim menggunakan satu molekul ATP untuk setiap molekul glukosa yang difosforilasi. Ini adalah reaksi ireversibel dan tergantung pada keberadaan ion magnesium (Mg2 +):
Glukosa + ATP → Glukosa 6-fosfat + ADP
2- Glukosa 6-fosfat yang dihasilkan diisomerisasi menjadi fruktosa 6-fosfat berkat kerja enzim fosfoglukosa isomerase (PGI). Ini adalah reaksi reversibel dan tidak melibatkan pengeluaran energi tambahan:
Glukosa 6-fosfat → Fruktosa 6-fosfat
3- Selanjutnya, langkah inversi energi lain melibatkan fosforilasi fruktosa 6-fosfat untuk membentuk fruktosa 1,6-bifosfat. Reaksi ini dikatalisis oleh enzim fosfofruktokinase-1 (PFK-1). Seperti langkah pertama dalam jalur, molekul donor gugus fosfat adalah ATP dan juga merupakan reaksi ireversibel.
Fruktosa 6-fosfat + ATP → Fruktosa 1,6-bifosfat + ADP
4- Dalam langkah glikolisis ini, ada pemecahan katalitik fruktosa 1,6-bifosfat menjadi dihidroksiaseton fosfat (DHAP) suatu ketosa, dan gliseraldehida 3-fosfat (GAP) suatu alsin. Kondensasi aldol ini dikatalisis oleh enzim aldolase dan merupakan proses reversibel.
Fruktosa 1,6-bifosfat → Dihidroksiaseton fosfat + gliseraldehida 3-fosfat
5- Reaksi terakhir dari fase inversi energi terdiri dari interkonversi triosa fosfat DHAP dan GAP yang dikatalisis oleh enzim triosa fosfat isomerase (TIM), fakta yang tidak memerlukan asupan energi tambahan dan juga merupakan proses reversibel.
Dihidroksiaseton fosfat Gliseraldehida 3-fosfat
– Fase perolehan energi
6- Gliseraldehida 3-fosfat digunakan “hilir” dalam jalur glikolitik sebagai substrat untuk reaksi oksidasi dan reaksi fosforilasi, dikatalisis oleh enzim yang sama, gliseraldehida 3-fosfat dehidrogenase (GAPDH).
Enzim mengkatalisis oksidasi karbon C1 molekul menjadi asam karboksilat dan fosforilasinya pada posisi yang sama, menghasilkan 1,3-bisfosfogliserat. Selama reaksi, 2 molekul NAD + direduksi untuk setiap molekul glukosa dan 2 molekul fosfat anorganik digunakan.
2Gliseraldehida 3-fosfat + 2NAD + + 2Pi → 2 (1,3-bifosfogliserat) + 2NADH + 2H
Pada organisme aerobik, setiap NADH yang diproduksi dengan cara ini melewati rantai transpor elektron untuk berfungsi sebagai substrat untuk sintesis 6 molekul ATP melalui fosforilasi oksidatif.
7- Ini adalah langkah sintesis ATP pertama dalam glikolisis dan melibatkan aksi fosfogliserat kinase (PGK) pada 1,3-bisfosfogliserat, mentransfer gugus fosforil (fosforilasi tingkat substrat) dari molekul ini ke molekul ADP, menghasilkan 2ATP dan 2 molekul 3-fosfogliserat (3PG) untuk setiap molekul glukosa.
2 (1,3-bisfosfogliserat) + 2ADP → 2 (3-fosfogliserat) + 2ATP
8- 3-fosfogliserat berfungsi sebagai substrat untuk enzim fosfogliserat mutase (PGM), yang mengubahnya menjadi 2-fosfogliserat dengan perpindahan gugus fosforil dari karbon 3 ke karbon 2 melalui reaksi dua langkah yang reversibel dan bergantung pada magnesium ion (Mg + 2).
2 (3-fosfogliserat) → 2 (2-fosfogliserat)
9- Enzim enolase mendehidrasi 2-fosfogliserat dan menghasilkan fosfoenolpiruvat (PEP) melalui reaksi yang tidak memerlukan penambahan energi tambahan dan bertujuan untuk menghasilkan senyawa berenergi tinggi, yang mampu menyumbangkan gugus fosforilnya dalam reaksi berikut.
2 (2-fosfogliserat) → 2fosfoenolpiruvat
10- Fosfoenolpiruvat adalah substrat untuk enzim piruvat kinase (PYK), yang bertanggung jawab untuk transfer gugus fosforil dalam molekul ini ke molekul ADP, sehingga mengkatalisis reaksi fosforilasi lain pada tingkat substrat.
Dalam reaksi, 2ATP dan 2 molekul piruvat diproduksi untuk setiap glukosa dan keberadaan kalium dan magnesium dalam bentuk ionik diperlukan.
2Fosfoenolpiruvat + 2ADP → 2Piruvat + 2ATP
Hasil bersih glikolisis, dengan cara ini, terdiri dari 2ATP dan 2NAD+ untuk setiap molekul glukosa yang memasuki jalur tersebut.
Jika sel dengan metabolisme aerobik, maka degradasi total molekul glukosa menghasilkan antara 30 dan 32 ATP melalui siklus Krebs dan rantai transpor elektron.
Produk dari glikolisis
Reaksi umum glikolisis adalah sebagai berikut:
Glukosa + 2NAD + + 2ADP + 2Pi → 2Piruvat + 2ATP + 2NADH + 2H +
Oleh karena itu, jika dianalisis secara singkat, dapat dipastikan bahwa produk utama dari jalur glikolitik adalah piruvat, ATP, NADH dan H.
Namun, nasib metabolisme masing-masing zat antara reaksi tergantung, sebagian besar, pada kebutuhan seluler, itulah sebabnya semua zat antara dapat dianggap sebagai produk reaksi, dan mereka dapat didaftar sebagai berikut:
– Glukosa 6-fosfat
– Fruktosa 6-fosfat
– Fruktosa 1,6-bifosfat
– Dihidroksiaseton fosfat dan gliseraldehida 3-fosfat
– 1,3-bifosfogliserat
– 3-fosfogliserat dan 2-fosfogliserat
– Fosfoenolpiruvat dan piruvat
Pentingnya
Terlepas dari kenyataan bahwa glikolisis, dengan sendirinya (orang dapat berbicara tentang glikolisis anaerobik), hanya menghasilkan sekitar 5% dari ATP yang dapat diekstraksi dari katabolisme aerobik glukosa, jalur metabolisme ini penting karena beberapa alasan:
– Ini berfungsi sebagai sumber energi “cepat”, terutama dalam situasi di mana hewan harus keluar dari keadaan istirahat dengan cepat, di mana proses oksidasi aerobik tidak akan cukup cepat.
– Serat otot rangka “putih” dalam tubuh manusia, misalnya, adalah serat berkedut cepat dan bergantung pada glikolisis anaerobik untuk berfungsi.
– Ketika, untuk beberapa alasan, sel perlu melakukan tanpa beberapa mitokondrianya (yang merupakan organel yang melakukan fosforilasi oksidatif bagian dari produk glikolitik, antara lain) sel menjadi lebih bergantung pada energi yang diperoleh melalui jalur glikolitik .
– Banyak sel bergantung pada glukosa sebagai sumber energi melalui jalur glikolitik, di antaranya sel darah merah , tidak memiliki organel internal, dan sel mata (terutama kornea) yang tidak memiliki kepadatan tinggi mitokondria menonjol .
Referensi
- Canback, B., Andersson, SGE, & Kurland, CG (2002). Filogeni global enzim glikolitik. Prosiding National Academy of Sciences, 99 (9), 6097-6102.
- Chaudhry R, Varacallo M. Biokimia, Glikolisis. [Diperbarui 2019 Apr 21]. Di: StatPearls [Internet]. Treasure Island (FL): Penerbitan StatPearls; 2020 Jan-. Tersedia dari: https://www.ncbi.nlm.nih.gov/books/NBK482303/
- Fothergill-Gilmore, LA, & Michels, PA (1993). Evolusi glikolisis. Kemajuan dalam biofisika dan biologi molekuler, 59 (2), 105-235.
- Kim, JW, & Dang, CV (2005). Peran multifaset dari enzim glikolitik. Tren dalam ilmu biokimia, 30 (3), 142-150.
- Kumari, A. (2017). Biokimia Manis: Mengingat Struktur, Siklus, dan Jalur oleh Mnemonik. Pers Akademik.
- Li, XB, Gu, JD, & Zhou, QH (2015). Tinjauan glikolisis aerobik dan enzim kuncinya – target baru untuk terapi kanker paru-paru. Kanker dada, 6 (1), 17-24.
