
Autophagy adalah sistem degradasi intraseluler terjadi sehingga diawetkan dalam lisosom dari semua sel eukariotik (dan vakuola ragi). Kata tersebut umumnya digunakan untuk merujuk pada degradasi komponen sitosol atau “bagian” sel yang “usang” atau yang telah berhenti bekerja dengan baik.
Istilah autophagy diciptakan pada tahun 1963 di Universitas Rockefeller oleh de Duve, yang juga mengamati dan menggambarkan proses endositosis seluler. Secara harfiah, kata autophagy berarti “mengkonsumsi diri sendiri”, meskipun beberapa penulis menggambarkannya sebagai “kanibalisme diri”.
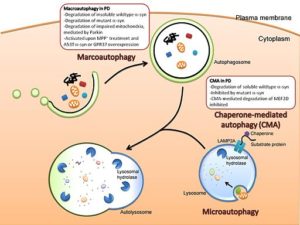
Representasi grafis dari Macroautophagy dan Microautophagy (Sumber: Cheung dan Ip [CC BY 3.0 (https://creativecommons.org/licenses/by/3.0)] melalui Wikimedia Commons)
Sistem ini berbeda dari degradasi yang dimediasi proteasom dalam autophagy yang mampu menghilangkan organel intraseluler lengkap dan kompleks atau agregat protein besar secara non-selektif.
Terlepas dari fagositosis non-selektif ini, penyelidikan yang berbeda telah menunjukkan bahwa autophagy memiliki banyak implikasi fisiologis dan patologis. Karena diaktifkan selama periode adaptasi terhadap kelaparan, selama perkembangan, untuk eliminasi mikroorganisme yang menyerang, selama kematian sel terprogram, untuk eliminasi tumor, presentasi antigen, dll.
Indeks artikel
Karakteristik
Autophagy, seperti yang dibahas, adalah proses yang dimediasi oleh organel sitoplasma yang dikenal sebagai lisosom.
Proses “autophagy” dimulai dengan enkapsulasi organel yang akan didegradasi oleh membran ganda, membentuk badan membran yang dikenal sebagai autophagosome. Membran autofagosom selanjutnya menyatu dengan membran lisosom atau dengan endosom akhir.
Masing-masing langkah antara penyerapan, degradasi, dan pelepasan asam amino atau komponen lain untuk daur ulang ini memberikan fungsi yang berbeda dalam konteks seluler yang berbeda, menjadikan autophagy sebagai sistem yang sangat multifungsi.
Autophagy adalah proses yang cukup terkontrol, karena hanya komponen seluler yang ditandai yang diarahkan ke jalur degradasi ini dan penandaan umumnya terjadi selama proses recaraling seluler.
Misalnya, ketika sel hati membentuk respons detoksifikasi sebagai respons terhadap obat yang larut dalam lemak, retikulum endoplasma halusnya berproliferasi secara signifikan, dan ketika stimulus yang dihasilkan oleh obat berkurang, retikulum endoplasma halus yang berlebih dikeluarkan dari ruang sitosol melalui autofagi.
Induksi autofagi
Salah satu peristiwa yang paling sering memicu proses autophagic adalah kelaparan.
Tergantung pada organisme yang dipertimbangkan, berbagai jenis nutrisi penting dapat memicu sistem “daur ulang” ini. Dalam ragi, misalnya, meskipun kekurangan karbon dalam asam amino dan asam nukleat tertentu dapat menyebabkan autophagy, kekurangan nitrogen adalah stimulus yang paling efisien, yang juga berlaku untuk sel tumbuhan.
Meskipun belum sepenuhnya dipahami, sel memiliki “sensor” khusus untuk menentukan kapan nutrisi atau asam amino esensial berada dalam kondisi sangat rendah, dan dengan demikian memicu seluruh proses daur ulang melalui lisosom.
Pada mamalia, beberapa hormon berpartisipasi dalam regulasi (positif atau negatif) autophagy dalam sel-sel milik organ tertentu, seperti insulin, beberapa faktor pertumbuhan atau interleukin, dll.
Jenis
Ada tiga jenis utama autophagy di antara eukariota: autophagy makro, autophagy mikro, dan autophagy yang dimediasi pendamping. Kecuali ditentukan, istilah autophagy mengacu pada autophagy makro.
Meskipun ketiga jenis autophagy secara morfologis berbeda, semuanya berakhir pada pengangkutan zat ke lisosom untuk degradasi dan daur ulang.
Makroautofagi
Ini adalah jenis autophagy yang bergantung pada pembentukan de novo dari vesikel fagosit yang dikenal sebagai autophagosomes. Pembentukan vesikel ini tidak tergantung pada pembentukan “tunas” membran, karena mereka dibentuk oleh ekspansi.
Dalam ragi, pembentukan autofagosom dimulai di tempat tertentu yang dikenal sebagai PAS, sementara pada mamalia banyak tempat berbeda terjadi di sitosol, mungkin terkait dengan retikulum endoplasma melalui struktur yang dikenal sebagai “omegasom”.
Ukuran autofagosom sangat bervariasi dan tergantung pada organisme dan jenis molekul atau organel yang difagositosis. Diameternya dapat bervariasi dari 0,4-0,9 m pada ragi hingga 0,5-1,5 m pada mamalia.
Ketika membran autophagosome dan lisosom menyatu, isinya bercampur dan saat itulah pencernaan substrat target autophagy dimulai. Organel ini kemudian dikenal sebagai autolisosom.
Untuk beberapa penulis, macroautophagy dapat disubklasifikasikan, pada gilirannya, menjadi autophagy yang diinduksi dan autophagy dasar. Makroautofagi terinduksi digunakan untuk memproduksi asam amino setelah periode kelaparan yang berkepanjangan.
Makroautofagi basal mengacu pada mekanisme konstitutif (yang selalu aktif) penting untuk pergantian berbagai komponen sitosol dan organel intraseluler.
Mikroautofagi
Jenis autophagy ini mengacu pada proses di mana konten sitoplasma diperkenalkan ke lisosom melalui invaginasi yang terjadi pada membran organel tersebut.
Setelah dimasukkan ke dalam lisosom, vesikel yang dihasilkan oleh invaginasi ini mengapung bebas di lumen sampai mereka dilisiskan dan isinya dilepaskan dan didegradasi oleh enzim tertentu.
Autophagy yang dimediasi pendamping
Jenis autophagy ini hanya dilaporkan untuk sel mamalia. Tidak seperti macroautophagy dan microautophagy, di mana beberapa bagian sitosol difagositosis secara non-spesifik, autophagy yang dimediasi oleh pendamping cukup spesifik, karena bergantung pada keberadaan urutan pentapeptida tertentu dalam substrat yang akan difagositosis.
Beberapa peneliti telah menentukan bahwa motif pentapeptida ini terkait dengan urutan KFERQ dan ditemukan di lebih dari 30% protein sitosol.
Ini disebut “dimediasi pendamping” karena protein pendamping bertanggung jawab untuk menjaga motif yang dilestarikan ini tetap terbuka untuk memfasilitasi pengenalannya dan mencegah protein terlipat di atasnya.
Protein dengan tag ini ditranslokasi ke lumen lisosom dan di sana mereka terdegradasi. Banyak substrat untuk degradasi adalah enzim glikolitik, faktor transkripsi dan penghambatnya, protein pengikat kalsium atau lipid, subunit proteasome, dan beberapa protein yang terlibat dalam perdagangan vesikular.
Seperti dua jenis autophagy lainnya, autophagy yang dimediasi pendamping adalah proses yang diatur di banyak tingkatan, mulai dari pengenalan label hingga transportasi dan degradasi substrat dalam lisosom.
Fitur
Salah satu fungsi utama dari proses autophagic adalah penghapusan organel tua atau “basi”, yang ditandai oleh berbagai rute untuk degradasi dalam lisosom.
Berkat pengamatan mikrofotograf elektron lisosom dalam sel mamalia, keberadaan peroksisom dan mitokondria telah terdeteksi di dalamnya .
Dalam sel hati, misalnya, waktu hidup rata-rata mitokondria adalah 10 hari, setelah itu organel ini difagositosis oleh lisosom, di mana ia didegradasi dan komponennya didaur ulang untuk tujuan metabolisme yang berbeda.
Dalam kondisi konsentrasi nutrisi yang rendah, sel dapat memicu pembentukan autofagosom untuk secara selektif “menangkap” bagian sitosol, serta metabolit yang dicerna dalam autofagosom ini dapat membantu sel untuk bertahan hidup ketika kondisi eksternal membatasi dari sudut pandang. dari segi nutrisi.
Peran dalam kesehatan dan pembangunan
Autophagy memiliki fungsi penting dalam restrukturisasi sel dalam proses diferensiasi, karena berpartisipasi dalam membuang bagian sitosol yang tidak diperlukan pada waktu tertentu.
Ini juga memiliki implikasi penting bagi kesehatan sel, karena merupakan bagian dari mekanisme pertahanan terhadap virus dan bakteri yang menyerang.
Studi Yoshinori Ohsumi
Yoshinori Ohsumi, seorang peneliti Jepang pemenang Hadiah Nobel 2016 dalam Fisiologi dan Kedokteran, menggambarkan mekanisme molekuler autophagy dalam ragi sambil mempelajari nasib metabolisme banyak protein dan vakuola organisme bersel tunggal ini.
Dalam karyanya, Ohsumi tidak hanya mengidentifikasi protein dan jalur yang terlibat dalam proses tersebut, tetapi juga menunjukkan bagaimana jalur autophagy diatur berkat aksi protein yang mampu “merasakan” status metabolisme yang berbeda.
Pekerjaan mereka dimulai dengan pengamatan mikroskopis yang tepat dari vakuola selama peristiwa degradasi yang intens. Vakuola dianggap sebagai tempat penyimpanan “sampah” ragi dan puing-puing seluler.
Dengan mengamati ragi dengan genotipe mutan yang cacat untuk gen berbeda yang terkait atau secara hipotetis terkait dengan autophagy (dikenal sebagai gen ATG ), peneliti ini dan rekan-rekannya mampu menggambarkan sistem autophagic ragi pada tingkat genetik.
Selanjutnya, kelompok peneliti ini menentukan karakteristik genetik utama dari protein yang dikodekan oleh gen-gen ini dan memberikan kontribusi signifikan tentang interaksi mereka dan pembentukan kompleks yang bertanggung jawab untuk inisiasi dan pelaksanaan autophagy dalam ragi.
Berkat karya Yoshinori Ohsumi, hari ini kita lebih memahami aspek molekuler autophagy, serta implikasi pentingnya dalam berfungsinya sel dan organ yang membentuk kita dengan benar.
Referensi
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2015). Biologi Molekuler Sel (edisi ke-6). New York: Ilmu Garland.
- Klionsky, DJ, & Emr, SD (2000). Autophagy sebagai jalur degradasi seluler yang diatur. Sains , 290 , 1717-1721.
- Mizushima, N. (2007). Autophagy: proses dan fungsi. Gen & Perkembangan , 21 , 2861–2873.
- Mizushima, Noboru, & Komatsu, M. (2011). Autophagy: Renovasi sel dan jaringan. Sel , 147 , 728-741.
- Rabinowitz, JD, & White, E. (2010). Autofagi dan metabolisme. Sains , 330 , 1344-1348.
