dihibridismo , genetik, mendefinisikan studi simultan dari dua karakteristik keturunan yang berbeda, dan dengan perpanjangan, dari mereka yang ekspresi tergantung pada dua gen yang berbeda bahkan jika itu adalah dari karakter yang sama
Tujuh sifat yang dianalisis Mendel berguna baginya dalam mengajukan teorinya tentang pewarisan sifat karena, antara lain, gen yang bertanggung jawab atas manifestasinya memiliki alel yang kontras yang fenotipenya mudah dianalisis, dan karena masing-masing menentukan ekspresi dari satu karakter.
Artinya, mereka adalah sifat-sifat monogenik yang kondisi hibridnya (monohibrid) memungkinkan menentukan hubungan dominasi / resesif antara alel dari gen tunggal itu.
Ketika Mendel menganalisis warisan bersama dari dua karakter yang berbeda, dia melanjutkan seperti yang dia lakukan dengan karakter tunggal. Dia memperoleh hibrida ganda (dihibrida) yang memungkinkan dia untuk memeriksa:
- Bahwa masing-masing mematuhi segregasi independen yang dia amati pada persilangan monohibrid.
- Selanjutnya, dalam persilangan dihibrida, manifestasi masing-masing karakter tidak tergantung pada manifestasi fenotipik yang lain. Artinya, faktor warisan mereka, apa pun itu, didistribusikan secara independen.
Kita sekarang tahu bahwa pewarisan karakter sedikit lebih kompleks daripada yang diamati Mendel, tetapi juga bahwa pada dasarnya Mendel sepenuhnya benar.
Perkembangan genetika selanjutnya memungkinkan untuk menunjukkan bahwa persilangan dihibrida dan analisisnya (dihibridisme), seperti yang awalnya dapat ditunjukkan oleh Bateson, dapat menjadi sumber penemuan yang tiada habisnya dalam sains abad ke-20 yang kuat dan baru lahir ini.
Melalui penggunaan yang cerdas, mereka dapat memberikan gambaran yang lebih jelas kepada ahli genetika tentang perilaku dan sifat gen.
Indeks artikel
Persilangan dihibrid dengan karakter yang berbeda
Jika kita menganalisis produk dari persilangan monohibrid Aa X Aa , kita dapat melihat bahwa itu sama dengan mengembangkan produk yang luar biasa ( A + a ) 2 = AA + 2Aa + aa .
Ekspresi di sebelah kiri mencakup dua jenis gamet yang dapat menghasilkan salah satu parental heterozigot untuk gen A / a ; dengan mengkuadratkan kita menunjukkan bahwa kedua orang tua memiliki konstitusi yang identik untuk gen yang diteliti [yaitu, itu adalah persilangan monohibrid ( A + a ) X ( A + a )].
Ekspresi di sebelah kanan memberi kita genotipe (dan oleh karena itu fenotipe disimpulkan) dan proporsi yang diharapkan berasal dari persilangan.
Oleh karena itu, kita dapat mengamati secara langsung proporsi genotip yang diturunkan dari hukum pertama (1: 2: 1), serta proporsi fenotipik yang dijelaskan olehnya (1 AA +2 Aa = 3 A _ untuk setiap 1 aa , atau rasio fenotipik 3 :satu).
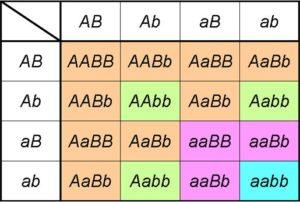
Jika sekarang kita mempertimbangkan persilangan untuk menganalisis pewarisan gen B , ekspresi dan proporsinya akan sama; pada kenyataannya, hal itu akan terjadi pada gen apa pun. Oleh karena itu, dalam persilangan dihibrida, kita sebenarnya memiliki pengembangan produk dari ( A + a ) 2 X ( B + b ) 2 .
Atau apa sama, jika persilangan dihibrida melibatkan dua gen yang berpartisipasi dalam pewarisan dua karakter yang tidak berhubungan, proporsi fenotipnya akan seperti yang diprediksi oleh hukum kedua: (3 A _: 1 aa ) X (3 B _: 1 bb ) = 9 A _ B _: 3 A _ bb : 3 aaB _: 1 aabb ).
Ini, tentu saja, diturunkan dari rasio genotipe terurut 4: 2: 2: 2: 2: 1: 1: 1: 1 yang dihasilkan dari produk ( A + a ) 2 X ( B + b ) 2 = ( AA + 2Aa + aa ) X ( BB + 2 Bb + bb ).
Kita mengundang Anda untuk memeriksanya sendiri untuk menganalisis sekarang apa yang terjadi ketika rasio fenotipik 9: 3: 3: 1 dari persilangan dihibrida “menyimpang” dari hubungan matematis yang jelas dan dapat diprediksi ini yang menjelaskan pewarisan independen dari dua karakter yang disandikan .oleh gen yang berbeda.
Manifestasi fenotip alternatif persilangan dihibrid
Ada dua cara utama persilangan dihibrid menyimpang dari apa yang “diharapkan”. Yang pertama adalah yang di mana kita menganalisis pewarisan bersama dari dua karakter yang berbeda, tetapi proporsi fenotipik yang diamati pada keturunannya memberikan dominasi yang jelas pada manifestasi fenotipe induk.
Kemungkinan besar itu adalah kasus gen terkait. Artinya, dua gen yang dianalisis, meskipun mereka berada di lokus yang berbeda, sangat dekat satu sama lain secara fisik sehingga mereka cenderung diwariskan bersama-sama dan, jelas, mereka tidak didistribusikan secara independen.
Keadaan lain, yang juga cukup umum, berasal dari fakta bahwa sebagian kecil dari sifat-sifat herediter adalah monogenik.
Sebaliknya, lebih dari dua gen terlibat dalam manifestasi sebagian besar sifat yang diwariskan.
Untuk alasan ini, selalu mungkin bahwa interaksi genetik yang terbentuk antara gen yang berpartisipasi dalam manifestasi suatu sifat tunggal bersifat kompleks dan melampaui hubungan dominasi atau resesif yang sederhana seperti yang diamati dalam hubungan alelik yang khas dari sifat monogenik.
Misalnya, manifestasi suatu sifat dapat melibatkan sekitar empat enzim dalam urutan tertentu untuk menghasilkan produk akhir yang bertanggung jawab atas manifestasi fenotipik dari fenotipe tipe liar.
Analisis yang memungkinkan untuk mengidentifikasi jumlah gen dari berbagai lokus yang berpartisipasi dalam manifestasi sifat genetik, serta urutan tindakannya, disebut analisis epistasis dan mungkin yang paling sering mendefinisikan apa yang kita sebut analisis genetik dalam pengertian yang paling klasik.
Sedikit lagi epistasis
Di akhir posting ini, proporsi fenotipik yang diamati dalam kasus epistasis yang paling umum disajikan – dan ini hanya memperhitungkan persilangan dihibrida.
Dengan meningkatkan jumlah gen yang terlibat dalam manifestasi sifat yang sama, kompleksitas interaksi gen dan interpretasinya jelas meningkat.
Selanjutnya, yang pada gilirannya dapat diambil sebagai aturan emas untuk diagnosis interaksi epistatik yang benar, penampilan fenotipe baru yang tidak ada pada generasi induk dapat diverifikasi.
Akhirnya, selain memungkinkan kita untuk menganalisis penampilan fenotipe baru dan proporsinya, analisis epistasis juga memungkinkan kita untuk menentukan urutan hierarki di mana gen yang berbeda dan produknya harus bermanifestasi dalam jalur tertentu untuk menjelaskan fenotipe yang terkait dengannya. mereka.
Gen manifestasi paling dasar atau awal adalah epistatik di atas yang lain, karena tanpa produk atau tindakannya, misalnya, mereka yang berada di hilirnya tidak akan dapat mengekspresikan diri, yang karenanya menjadi hipostatik terhadapnya.
Gen / produk ketiga dalam hierarki akan menjadi hipostatik untuk dua yang pertama, dan epistatik untuk setiap yang tersisa di jalur ekspresi gen ini.
Referensi
- Bateson, W. (1909). Prinsip Keturunan Mendel. Pers Universitas Cambridge. Cambridge, Inggris Raya
- Brooker, RJ (2017). Genetika: Analisis dan Prinsip. Pendidikan Tinggi McGraw-Hill, New York, NY, AS.
- Cordell, H. (2002). Epistasis: Apa artinya, apa artinya, dan metode statistik untuk mendeteksinya pada manusia. Genetika Molekuler Manusia, 11: 2463–2468.
- Cukup baik, UW (1984) Genetika. WB Saunders Co. Ltd, Pkiladelphia, PA, AS.
- Griffiths, AJF, Wessler, R., Carroll, SB, Doebley, J. (2015). Sebuah Pengantar Analisis Genetika (11 th ed.). New York: WH Freeman, New York, NY, AS.